JP4127567B2 - グリア細胞分裂誘発因子とその調製及び使用 - Google Patents
グリア細胞分裂誘発因子とその調製及び使用 Download PDFInfo
- Publication number
- JP4127567B2 JP4127567B2 JP50265594A JP50265594A JP4127567B2 JP 4127567 B2 JP4127567 B2 JP 4127567B2 JP 50265594 A JP50265594 A JP 50265594A JP 50265594 A JP50265594 A JP 50265594A JP 4127567 B2 JP4127567 B2 JP 4127567B2
- Authority
- JP
- Japan
- Prior art keywords
- ggf
- sequence
- cells
- cell
- polypeptide
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
- 238000002360 preparation method Methods 0.000 title description 20
- 230000002518 glial effect Effects 0.000 title description 15
- 239000003226 mitogen Substances 0.000 title description 11
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 22
- 239000013598 vector Substances 0.000 claims abstract description 22
- 108020004707 nucleic acids Proteins 0.000 claims abstract 7
- 102000039446 nucleic acids Human genes 0.000 claims abstract 7
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 219
- 210000004027 cell Anatomy 0.000 claims description 177
- 108090000623 proteins and genes Proteins 0.000 claims description 173
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 142
- 229920001184 polypeptide Polymers 0.000 claims description 101
- 238000000034 method Methods 0.000 claims description 90
- 210000004498 neuroglial cell Anatomy 0.000 claims description 41
- 230000027455 binding Effects 0.000 claims description 24
- 239000000203 mixture Substances 0.000 claims description 20
- 239000003446 ligand Substances 0.000 claims description 16
- 230000014509 gene expression Effects 0.000 claims description 13
- 238000009472 formulation Methods 0.000 claims description 12
- 238000000338 in vitro Methods 0.000 claims description 11
- 238000012258 culturing Methods 0.000 claims description 7
- 239000003814 drug Substances 0.000 claims description 7
- 238000004519 manufacturing process Methods 0.000 claims description 6
- 239000000546 pharmaceutical excipient Substances 0.000 claims description 4
- 239000003085 diluting agent Substances 0.000 claims description 3
- 239000002552 dosage form Substances 0.000 claims description 2
- 125000003275 alpha amino acid group Chemical group 0.000 claims 1
- 108090000556 Neuregulin-1 Proteins 0.000 abstract description 121
- 102000048238 Neuregulin-1 Human genes 0.000 abstract description 113
- 102000004169 proteins and genes Human genes 0.000 description 130
- 235000018102 proteins Nutrition 0.000 description 127
- 210000004116 schwann cell Anatomy 0.000 description 103
- 241000283690 Bos taurus Species 0.000 description 96
- 150000001413 amino acids Chemical group 0.000 description 89
- 108020004414 DNA Proteins 0.000 description 76
- 108020004635 Complementary DNA Proteins 0.000 description 65
- 108091028043 Nucleic acid sequence Proteins 0.000 description 63
- 238000004458 analytical method Methods 0.000 description 60
- 238000010804 cDNA synthesis Methods 0.000 description 59
- 239000002299 complementary DNA Substances 0.000 description 59
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 58
- 230000002297 mitogenic effect Effects 0.000 description 54
- 239000000523 sample Substances 0.000 description 54
- 239000000047 product Substances 0.000 description 52
- 230000000694 effects Effects 0.000 description 48
- 239000003636 conditioned culture medium Substances 0.000 description 45
- DTQVDTLACAAQTR-UHFFFAOYSA-N Trifluoroacetic acid Chemical compound OC(=O)C(F)(F)F DTQVDTLACAAQTR-UHFFFAOYSA-N 0.000 description 38
- 239000002609 medium Substances 0.000 description 37
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 36
- 102000005962 receptors Human genes 0.000 description 34
- 108020003175 receptors Proteins 0.000 description 34
- 238000000746 purification Methods 0.000 description 33
- 229940024606 amino acid Drugs 0.000 description 31
- 238000006243 chemical reaction Methods 0.000 description 31
- 239000000463 material Substances 0.000 description 31
- 238000003752 polymerase chain reaction Methods 0.000 description 31
- 239000013615 primer Substances 0.000 description 31
- 235000001014 amino acid Nutrition 0.000 description 30
- 239000000499 gel Substances 0.000 description 30
- 239000011780 sodium chloride Substances 0.000 description 29
- 239000002773 nucleotide Substances 0.000 description 27
- 125000003729 nucleotide group Chemical group 0.000 description 27
- 241000700159 Rattus Species 0.000 description 25
- 101100226904 Unknown prokaryotic organism FCS1 gene Proteins 0.000 description 24
- 238000002474 experimental method Methods 0.000 description 24
- 239000000872 buffer Substances 0.000 description 23
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 22
- 238000009396 hybridization Methods 0.000 description 22
- WOVKYSAHUYNSMH-RRKCRQDMSA-N 5-bromodeoxyuridine Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(Br)=C1 WOVKYSAHUYNSMH-RRKCRQDMSA-N 0.000 description 21
- 101000856426 Mus musculus Cullin-7 Proteins 0.000 description 21
- 239000002953 phosphate buffered saline Substances 0.000 description 21
- DBMJMQXJHONAFJ-UHFFFAOYSA-M Sodium laurylsulphate Chemical compound [Na+].CCCCCCCCCCCCOS([O-])(=O)=O DBMJMQXJHONAFJ-UHFFFAOYSA-M 0.000 description 20
- 229910052739 hydrogen Inorganic materials 0.000 description 20
- 239000013543 active substance Substances 0.000 description 19
- 230000001817 pituitary effect Effects 0.000 description 19
- 239000012634 fragment Substances 0.000 description 17
- 239000003102 growth factor Substances 0.000 description 17
- 230000032823 cell division Effects 0.000 description 16
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 16
- 231100000673 dose–response relationship Toxicity 0.000 description 16
- 239000012528 membrane Substances 0.000 description 16
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 15
- 239000012071 phase Substances 0.000 description 15
- 238000004007 reversed phase HPLC Methods 0.000 description 15
- 238000012163 sequencing technique Methods 0.000 description 15
- 210000002966 serum Anatomy 0.000 description 15
- 239000002751 oligonucleotide probe Substances 0.000 description 14
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 14
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 13
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 13
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 13
- 238000010348 incorporation Methods 0.000 description 13
- 238000010186 staining Methods 0.000 description 13
- 108700024394 Exon Proteins 0.000 description 12
- OHCQJHSOBUTRHG-KGGHGJDLSA-N FORSKOLIN Chemical compound O=C([C@@]12O)C[C@](C)(C=C)O[C@]1(C)[C@@H](OC(=O)C)[C@@H](O)[C@@H]1[C@]2(C)[C@@H](O)CCC1(C)C OHCQJHSOBUTRHG-KGGHGJDLSA-N 0.000 description 12
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 12
- 238000004128 high performance liquid chromatography Methods 0.000 description 12
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 12
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 12
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 11
- 238000003556 assay Methods 0.000 description 11
- 238000010828 elution Methods 0.000 description 11
- 230000001605 fetal effect Effects 0.000 description 11
- 210000002950 fibroblast Anatomy 0.000 description 11
- 210000004379 membrane Anatomy 0.000 description 11
- 230000026731 phosphorylation Effects 0.000 description 11
- 238000006366 phosphorylation reaction Methods 0.000 description 11
- 239000000126 substance Substances 0.000 description 11
- 206010006187 Breast cancer Diseases 0.000 description 10
- 208000026310 Breast neoplasm Diseases 0.000 description 10
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 10
- 238000005341 cation exchange Methods 0.000 description 10
- 238000000926 separation method Methods 0.000 description 10
- 239000001488 sodium phosphate Substances 0.000 description 10
- 229910000162 sodium phosphate Inorganic materials 0.000 description 10
- 238000013519 translation Methods 0.000 description 10
- 230000014616 translation Effects 0.000 description 10
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 10
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 9
- 230000006820 DNA synthesis Effects 0.000 description 9
- 208000012902 Nervous system disease Diseases 0.000 description 9
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 9
- 238000005119 centrifugation Methods 0.000 description 9
- 238000004587 chromatography analysis Methods 0.000 description 9
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 9
- 239000012091 fetal bovine serum Substances 0.000 description 9
- 238000001502 gel electrophoresis Methods 0.000 description 9
- 108020004999 messenger RNA Proteins 0.000 description 9
- 230000036961 partial effect Effects 0.000 description 9
- 230000008488 polyadenylation Effects 0.000 description 9
- 230000002441 reversible effect Effects 0.000 description 9
- 239000000243 solution Substances 0.000 description 9
- 241000894007 species Species 0.000 description 9
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 8
- QCVGEOXPDFCNHA-UHFFFAOYSA-N 5,5-dimethyl-2,4-dioxo-1,3-oxazolidine-3-carboxamide Chemical compound CC1(C)OC(=O)N(C(N)=O)C1=O QCVGEOXPDFCNHA-UHFFFAOYSA-N 0.000 description 8
- 239000003298 DNA probe Substances 0.000 description 8
- 108010000912 Egg Proteins Proteins 0.000 description 8
- 102000002322 Egg Proteins Human genes 0.000 description 8
- 206010028980 Neoplasm Diseases 0.000 description 8
- 108020005187 Oligonucleotide Probes Proteins 0.000 description 8
- 108091000080 Phosphotransferase Proteins 0.000 description 8
- 238000001516 cell proliferation assay Methods 0.000 description 8
- 239000003795 chemical substances by application Substances 0.000 description 8
- 235000014103 egg white Nutrition 0.000 description 8
- 238000002523 gelfiltration Methods 0.000 description 8
- 229910052588 hydroxylapatite Inorganic materials 0.000 description 8
- XYJRXVWERLGGKC-UHFFFAOYSA-D pentacalcium;hydroxide;triphosphate Chemical compound [OH-].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O XYJRXVWERLGGKC-UHFFFAOYSA-D 0.000 description 8
- 102000020233 phosphotransferase Human genes 0.000 description 8
- 230000004936 stimulating effect Effects 0.000 description 8
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 8
- 238000001262 western blot Methods 0.000 description 8
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 7
- 241000238631 Hexapoda Species 0.000 description 7
- 102400000058 Neuregulin-1 Human genes 0.000 description 7
- 238000012408 PCR amplification Methods 0.000 description 7
- 230000004071 biological effect Effects 0.000 description 7
- 230000015572 biosynthetic process Effects 0.000 description 7
- 210000004556 brain Anatomy 0.000 description 7
- 230000004663 cell proliferation Effects 0.000 description 7
- 238000010367 cloning Methods 0.000 description 7
- 238000010276 construction Methods 0.000 description 7
- 230000006378 damage Effects 0.000 description 7
- 238000001962 electrophoresis Methods 0.000 description 7
- 238000002955 isolation Methods 0.000 description 7
- 210000004126 nerve fiber Anatomy 0.000 description 7
- 238000003199 nucleic acid amplification method Methods 0.000 description 7
- 239000013612 plasmid Substances 0.000 description 7
- 230000004044 response Effects 0.000 description 7
- 238000012216 screening Methods 0.000 description 7
- 239000012679 serum free medium Substances 0.000 description 7
- 229910052709 silver Inorganic materials 0.000 description 7
- 239000004332 silver Substances 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- 108020003215 DNA Probes Proteins 0.000 description 6
- SUZLHDUTVMZSEV-UHFFFAOYSA-N Deoxycoleonol Natural products C12C(=O)CC(C)(C=C)OC2(C)C(OC(=O)C)C(O)C2C1(C)C(O)CCC2(C)C SUZLHDUTVMZSEV-UHFFFAOYSA-N 0.000 description 6
- 108091034117 Oligonucleotide Proteins 0.000 description 6
- 229920002684 Sepharose Polymers 0.000 description 6
- 230000003321 amplification Effects 0.000 description 6
- 229940098773 bovine serum albumin Drugs 0.000 description 6
- 238000004113 cell culture Methods 0.000 description 6
- 239000013592 cell lysate Substances 0.000 description 6
- 239000013553 cell monolayer Substances 0.000 description 6
- 210000003855 cell nucleus Anatomy 0.000 description 6
- 229920002678 cellulose Polymers 0.000 description 6
- 239000001913 cellulose Substances 0.000 description 6
- 210000000991 chicken egg Anatomy 0.000 description 6
- OHCQJHSOBUTRHG-UHFFFAOYSA-N colforsin Natural products OC12C(=O)CC(C)(C=C)OC1(C)C(OC(=O)C)C(O)C1C2(C)C(O)CCC1(C)C OHCQJHSOBUTRHG-UHFFFAOYSA-N 0.000 description 6
- 235000018417 cysteine Nutrition 0.000 description 6
- 230000013595 glycosylation Effects 0.000 description 6
- 238000006206 glycosylation reaction Methods 0.000 description 6
- 239000001963 growth medium Substances 0.000 description 6
- 238000011534 incubation Methods 0.000 description 6
- 230000008747 mitogenic response Effects 0.000 description 6
- 210000003007 myelin sheath Anatomy 0.000 description 6
- 210000005036 nerve Anatomy 0.000 description 6
- 229920002401 polyacrylamide Polymers 0.000 description 6
- 230000035755 proliferation Effects 0.000 description 6
- BDERNNFJNOPAEC-UHFFFAOYSA-N propan-1-ol Chemical compound CCCO BDERNNFJNOPAEC-UHFFFAOYSA-N 0.000 description 6
- 108091008146 restriction endonucleases Proteins 0.000 description 6
- 239000011734 sodium Substances 0.000 description 6
- 230000000638 stimulation Effects 0.000 description 6
- 238000003786 synthesis reaction Methods 0.000 description 6
- 210000001519 tissue Anatomy 0.000 description 6
- 238000005406 washing Methods 0.000 description 6
- 229920002134 Carboxymethyl cellulose Polymers 0.000 description 5
- 108091026890 Coding region Proteins 0.000 description 5
- 101710081103 Cuticular glutathione peroxidase Proteins 0.000 description 5
- 102000004190 Enzymes Human genes 0.000 description 5
- 108090000790 Enzymes Proteins 0.000 description 5
- 208000032612 Glial tumor Diseases 0.000 description 5
- 206010018338 Glioma Diseases 0.000 description 5
- 239000000020 Nitrocellulose Substances 0.000 description 5
- 108010038512 Platelet-Derived Growth Factor Proteins 0.000 description 5
- 102000010780 Platelet-Derived Growth Factor Human genes 0.000 description 5
- 239000012980 RPMI-1640 medium Substances 0.000 description 5
- 229920005654 Sephadex Polymers 0.000 description 5
- 239000012507 Sephadex™ Substances 0.000 description 5
- 238000002105 Southern blotting Methods 0.000 description 5
- 239000012298 atmosphere Substances 0.000 description 5
- 230000010261 cell growth Effects 0.000 description 5
- 210000003169 central nervous system Anatomy 0.000 description 5
- 230000008859 change Effects 0.000 description 5
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 5
- 230000029087 digestion Effects 0.000 description 5
- 229940088598 enzyme Drugs 0.000 description 5
- 239000000284 extract Substances 0.000 description 5
- 238000001914 filtration Methods 0.000 description 5
- 238000003119 immunoblot Methods 0.000 description 5
- 238000012986 modification Methods 0.000 description 5
- 210000003205 muscle Anatomy 0.000 description 5
- 210000002569 neuron Anatomy 0.000 description 5
- 229920001220 nitrocellulos Polymers 0.000 description 5
- 238000007911 parenteral administration Methods 0.000 description 5
- 239000008188 pellet Substances 0.000 description 5
- 230000008569 process Effects 0.000 description 5
- 230000002062 proliferating effect Effects 0.000 description 5
- 230000002829 reductive effect Effects 0.000 description 5
- 230000008929 regeneration Effects 0.000 description 5
- 238000011069 regeneration method Methods 0.000 description 5
- 238000010561 standard procedure Methods 0.000 description 5
- 238000012360 testing method Methods 0.000 description 5
- 238000013518 transcription Methods 0.000 description 5
- 238000000108 ultra-filtration Methods 0.000 description 5
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 4
- 108010009685 Cholinergic Receptors Proteins 0.000 description 4
- 108020004705 Codon Proteins 0.000 description 4
- 102100032029 Epidermal growth factor-like protein 6 Human genes 0.000 description 4
- ZHNUHDYFZUAESO-UHFFFAOYSA-N Formamide Chemical compound NC=O ZHNUHDYFZUAESO-UHFFFAOYSA-N 0.000 description 4
- 101000921196 Homo sapiens Epidermal growth factor-like protein 6 Proteins 0.000 description 4
- 101000851176 Homo sapiens Pro-epidermal growth factor Proteins 0.000 description 4
- 101001109800 Homo sapiens Pro-neuregulin-1, membrane-bound isoform Proteins 0.000 description 4
- 208000025966 Neurological disease Diseases 0.000 description 4
- 241000283973 Oryctolagus cuniculus Species 0.000 description 4
- 229930182555 Penicillin Natural products 0.000 description 4
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 4
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 4
- 238000012300 Sequence Analysis Methods 0.000 description 4
- 210000001744 T-lymphocyte Anatomy 0.000 description 4
- 239000007983 Tris buffer Substances 0.000 description 4
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 4
- 208000027418 Wounds and injury Diseases 0.000 description 4
- 102000034337 acetylcholine receptors Human genes 0.000 description 4
- 239000002253 acid Substances 0.000 description 4
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 4
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 4
- 235000011130 ammonium sulphate Nutrition 0.000 description 4
- 239000011324 bead Substances 0.000 description 4
- 230000000903 blocking effect Effects 0.000 description 4
- 238000009835 boiling Methods 0.000 description 4
- 239000001768 carboxy methyl cellulose Substances 0.000 description 4
- 239000008112 carboxymethyl-cellulose Substances 0.000 description 4
- 150000001875 compounds Chemical class 0.000 description 4
- 238000010790 dilution Methods 0.000 description 4
- 239000012895 dilution Substances 0.000 description 4
- 230000012010 growth Effects 0.000 description 4
- 102000055650 human NRG1 Human genes 0.000 description 4
- 238000000021 kinase assay Methods 0.000 description 4
- 238000002372 labelling Methods 0.000 description 4
- GVUGOAYIVIDWIO-UFWWTJHBSA-N nepidermin Chemical compound C([C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)NC(=O)CNC(=O)[C@@H](NC(=O)[C@@H](NC(=O)[C@H](CS)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CS)NC(=O)[C@H](C)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](C)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C(C)C)C(C)C)C1=CC=C(O)C=C1 GVUGOAYIVIDWIO-UFWWTJHBSA-N 0.000 description 4
- 210000000653 nervous system Anatomy 0.000 description 4
- 201000001119 neuropathy Diseases 0.000 description 4
- 230000007823 neuropathy Effects 0.000 description 4
- 229940049954 penicillin Drugs 0.000 description 4
- 210000000578 peripheral nerve Anatomy 0.000 description 4
- 239000013641 positive control Substances 0.000 description 4
- 210000001883 posterior pituitary gland Anatomy 0.000 description 4
- 229910052700 potassium Inorganic materials 0.000 description 4
- 238000011160 research Methods 0.000 description 4
- 150000003839 salts Chemical class 0.000 description 4
- 229960005322 streptomycin Drugs 0.000 description 4
- 230000001225 therapeutic effect Effects 0.000 description 4
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 4
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 description 4
- 239000003643 water by type Substances 0.000 description 4
- GEYOCULIXLDCMW-UHFFFAOYSA-N 1,2-phenylenediamine Chemical compound NC1=CC=CC=C1N GEYOCULIXLDCMW-UHFFFAOYSA-N 0.000 description 3
- IEQAICDLOKRSRL-UHFFFAOYSA-N 2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-[2-(2-dodecoxyethoxy)ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethoxy]ethanol Chemical compound CCCCCCCCCCCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCOCCO IEQAICDLOKRSRL-UHFFFAOYSA-N 0.000 description 3
- BFSVOASYOCHEOV-UHFFFAOYSA-N 2-diethylaminoethanol Chemical compound CCN(CC)CCO BFSVOASYOCHEOV-UHFFFAOYSA-N 0.000 description 3
- 229920000936 Agarose Polymers 0.000 description 3
- 102100038778 Amphiregulin Human genes 0.000 description 3
- 108010033760 Amphiregulin Proteins 0.000 description 3
- 108010039627 Aprotinin Proteins 0.000 description 3
- 102100035680 Cadherin EGF LAG seven-pass G-type receptor 2 Human genes 0.000 description 3
- 102100035671 Cadherin EGF LAG seven-pass G-type receptor 3 Human genes 0.000 description 3
- 102000003846 Carbonic anhydrases Human genes 0.000 description 3
- 108090000209 Carbonic anhydrases Proteins 0.000 description 3
- 102000014914 Carrier Proteins Human genes 0.000 description 3
- 108010035532 Collagen Proteins 0.000 description 3
- 102000008186 Collagen Human genes 0.000 description 3
- 238000001712 DNA sequencing Methods 0.000 description 3
- 208000016192 Demyelinating disease Diseases 0.000 description 3
- 206010012305 Demyelination Diseases 0.000 description 3
- 102000018233 Fibroblast Growth Factor Human genes 0.000 description 3
- 108050007372 Fibroblast Growth Factor Proteins 0.000 description 3
- 102000003971 Fibroblast Growth Factor 1 Human genes 0.000 description 3
- 108090000386 Fibroblast Growth Factor 1 Proteins 0.000 description 3
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 3
- 244000068988 Glycine max Species 0.000 description 3
- 235000010469 Glycine max Nutrition 0.000 description 3
- 108090000288 Glycoproteins Proteins 0.000 description 3
- 102000003886 Glycoproteins Human genes 0.000 description 3
- 101000715674 Homo sapiens Cadherin EGF LAG seven-pass G-type receptor 2 Proteins 0.000 description 3
- 101000715671 Homo sapiens Cadherin EGF LAG seven-pass G-type receptor 3 Proteins 0.000 description 3
- 101000955263 Homo sapiens Multiple epidermal growth factor-like domains protein 6 Proteins 0.000 description 3
- 101000955249 Homo sapiens Multiple epidermal growth factor-like domains protein 8 Proteins 0.000 description 3
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 3
- 108010053229 Lysyl endopeptidase Proteins 0.000 description 3
- 102100039005 Multiple epidermal growth factor-like domains protein 6 Human genes 0.000 description 3
- 102100038990 Multiple epidermal growth factor-like domains protein 8 Human genes 0.000 description 3
- 102000016943 Muramidase Human genes 0.000 description 3
- 108010014251 Muramidase Proteins 0.000 description 3
- 108010062010 N-Acetylmuramoyl-L-alanine Amidase Proteins 0.000 description 3
- 230000004988 N-glycosylation Effects 0.000 description 3
- 239000004677 Nylon Substances 0.000 description 3
- 108700020796 Oncogene Proteins 0.000 description 3
- 239000002033 PVDF binder Substances 0.000 description 3
- 102000000447 Peptide-N4-(N-acetyl-beta-glucosaminyl) Asparagine Amidase Human genes 0.000 description 3
- 108010055817 Peptide-N4-(N-acetyl-beta-glucosaminyl) Asparagine Amidase Proteins 0.000 description 3
- 108010065081 Phosphorylase b Proteins 0.000 description 3
- 108090000412 Protein-Tyrosine Kinases Proteins 0.000 description 3
- 102000004022 Protein-Tyrosine Kinases Human genes 0.000 description 3
- 101710100968 Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 3
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 3
- 102100038126 Tenascin Human genes 0.000 description 3
- 108010008125 Tenascin Proteins 0.000 description 3
- 229920004890 Triton X-100 Polymers 0.000 description 3
- 229940122618 Trypsin inhibitor Drugs 0.000 description 3
- 101710162629 Trypsin inhibitor Proteins 0.000 description 3
- 108010006886 Vitrogen Proteins 0.000 description 3
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 3
- SXEHKFHPFVVDIR-UHFFFAOYSA-N [4-(4-hydrazinylphenyl)phenyl]hydrazine Chemical compound C1=CC(NN)=CC=C1C1=CC=C(NN)C=C1 SXEHKFHPFVVDIR-UHFFFAOYSA-N 0.000 description 3
- 238000001042 affinity chromatography Methods 0.000 description 3
- 239000011543 agarose gel Substances 0.000 description 3
- 229960004405 aprotinin Drugs 0.000 description 3
- 210000001130 astrocyte Anatomy 0.000 description 3
- 238000000211 autoradiogram Methods 0.000 description 3
- 210000003050 axon Anatomy 0.000 description 3
- 108091008324 binding proteins Proteins 0.000 description 3
- 210000000481 breast Anatomy 0.000 description 3
- 150000001720 carbohydrates Chemical group 0.000 description 3
- 235000010948 carboxy methyl cellulose Nutrition 0.000 description 3
- 238000005277 cation exchange chromatography Methods 0.000 description 3
- 230000005779 cell damage Effects 0.000 description 3
- 208000037887 cell injury Diseases 0.000 description 3
- 229920001436 collagen Polymers 0.000 description 3
- 238000011161 development Methods 0.000 description 3
- 239000011544 gradient gel Substances 0.000 description 3
- 108010034429 heregulin alpha Proteins 0.000 description 3
- 238000001727 in vivo Methods 0.000 description 3
- 230000006698 induction Effects 0.000 description 3
- ZPNFWUPYTFPOJU-LPYSRVMUSA-N iniprol Chemical compound C([C@H]1C(=O)NCC(=O)NCC(=O)N[C@H]2CSSC[C@H]3C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@H](C(N[C@H](C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC=4C=CC=CC=4)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC=4C=CC=CC=4)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C)NC(=O)[C@H](CCCNC(N)=N)NC2=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC=2C=CC=CC=2)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H]2N(CCC2)C(=O)[C@@H](N)CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N2[C@@H](CCC2)C(=O)N2[C@@H](CCC2)C(=O)N[C@@H](CC=2C=CC(O)=CC=2)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N2[C@@H](CCC2)C(=O)N3)C(=O)NCC(=O)NCC(=O)N[C@@H](C)C(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@H](C(=O)N1)C(C)C)[C@@H](C)O)[C@@H](C)CC)=O)[C@@H](C)CC)C1=CC=C(O)C=C1 ZPNFWUPYTFPOJU-LPYSRVMUSA-N 0.000 description 3
- 208000014674 injury Diseases 0.000 description 3
- 210000003734 kidney Anatomy 0.000 description 3
- 238000004811 liquid chromatography Methods 0.000 description 3
- 239000006166 lysate Substances 0.000 description 3
- 239000004325 lysozyme Substances 0.000 description 3
- 229960000274 lysozyme Drugs 0.000 description 3
- 235000010335 lysozyme Nutrition 0.000 description 3
- 230000001404 mediated effect Effects 0.000 description 3
- MYWUZJCMWCOHBA-VIFPVBQESA-N methamphetamine Chemical compound CN[C@@H](C)CC1=CC=CC=C1 MYWUZJCMWCOHBA-VIFPVBQESA-N 0.000 description 3
- 230000004048 modification Effects 0.000 description 3
- 238000010369 molecular cloning Methods 0.000 description 3
- 229920001778 nylon Polymers 0.000 description 3
- 210000004248 oligodendroglia Anatomy 0.000 description 3
- 229920002981 polyvinylidene fluoride Polymers 0.000 description 3
- 239000012521 purified sample Substances 0.000 description 3
- 239000011535 reaction buffer Substances 0.000 description 3
- 230000008439 repair process Effects 0.000 description 3
- 239000011347 resin Substances 0.000 description 3
- 229920005989 resin Polymers 0.000 description 3
- 230000000717 retained effect Effects 0.000 description 3
- 210000003497 sciatic nerve Anatomy 0.000 description 3
- 230000028327 secretion Effects 0.000 description 3
- FQENQNTWSFEDLI-UHFFFAOYSA-J sodium diphosphate Chemical compound [Na+].[Na+].[Na+].[Na+].[O-]P([O-])(=O)OP([O-])([O-])=O FQENQNTWSFEDLI-UHFFFAOYSA-J 0.000 description 3
- 229940048086 sodium pyrophosphate Drugs 0.000 description 3
- 235000019818 tetrasodium diphosphate Nutrition 0.000 description 3
- 239000001577 tetrasodium phosphonato phosphate Substances 0.000 description 3
- 230000035897 transcription Effects 0.000 description 3
- 239000002753 trypsin inhibitor Substances 0.000 description 3
- 229940045145 uridine Drugs 0.000 description 3
- MXHRCPNRJAMMIM-SHYZEUOFSA-N 2'-deoxyuridine Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 MXHRCPNRJAMMIM-SHYZEUOFSA-N 0.000 description 2
- AXAVXPMQTGXXJZ-UHFFFAOYSA-N 2-aminoacetic acid;2-amino-2-(hydroxymethyl)propane-1,3-diol Chemical compound NCC(O)=O.OCC(N)(CO)CO AXAVXPMQTGXXJZ-UHFFFAOYSA-N 0.000 description 2
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 2
- 108010088751 Albumins Proteins 0.000 description 2
- 102000009027 Albumins Human genes 0.000 description 2
- NLXLAEXVIDQMFP-UHFFFAOYSA-N Ammonia chloride Chemical compound [NH4+].[Cl-] NLXLAEXVIDQMFP-UHFFFAOYSA-N 0.000 description 2
- ATRRKUHOCOJYRX-UHFFFAOYSA-N Ammonium bicarbonate Chemical compound [NH4+].OC([O-])=O ATRRKUHOCOJYRX-UHFFFAOYSA-N 0.000 description 2
- 229910000013 Ammonium bicarbonate Inorganic materials 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- HEDRZPFGACZZDS-UHFFFAOYSA-N Chloroform Chemical compound ClC(Cl)Cl HEDRZPFGACZZDS-UHFFFAOYSA-N 0.000 description 2
- 239000003155 DNA primer Substances 0.000 description 2
- 229920002307 Dextran Polymers 0.000 description 2
- 102000012545 EGF-like domains Human genes 0.000 description 2
- 108050002150 EGF-like domains Proteins 0.000 description 2
- 238000002965 ELISA Methods 0.000 description 2
- 101800003838 Epidermal growth factor Proteins 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- 108010007979 Glycocholic Acid Proteins 0.000 description 2
- ZRALSGWEFCBTJO-UHFFFAOYSA-N Guanidine Chemical compound NC(N)=N ZRALSGWEFCBTJO-UHFFFAOYSA-N 0.000 description 2
- 101000955254 Homo sapiens Multiple epidermal growth factor-like domains protein 9 Proteins 0.000 description 2
- 108090000144 Human Proteins Proteins 0.000 description 2
- 102000003839 Human Proteins Human genes 0.000 description 2
- 206010062767 Hypophysitis Diseases 0.000 description 2
- 108060003951 Immunoglobulin Proteins 0.000 description 2
- 108010002350 Interleukin-2 Proteins 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 2
- TWRXJAOTZQYOKJ-UHFFFAOYSA-L Magnesium chloride Chemical compound [Mg+2].[Cl-].[Cl-] TWRXJAOTZQYOKJ-UHFFFAOYSA-L 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 102100038989 Multiple epidermal growth factor-like domains protein 9 Human genes 0.000 description 2
- 208000028389 Nerve injury Diseases 0.000 description 2
- 229910019142 PO4 Inorganic materials 0.000 description 2
- OAICVXFJPJFONN-UHFFFAOYSA-N Phosphorus Chemical compound [P] OAICVXFJPJFONN-UHFFFAOYSA-N 0.000 description 2
- 108010021757 Polynucleotide 5'-Hydroxyl-Kinase Proteins 0.000 description 2
- 102000008422 Polynucleotide 5'-hydroxyl-kinase Human genes 0.000 description 2
- 102100033237 Pro-epidermal growth factor Human genes 0.000 description 2
- 239000013614 RNA sample Substances 0.000 description 2
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical compound [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 2
- QAOWNCQODCNURD-UHFFFAOYSA-N Sulfuric acid Chemical compound OS(O)(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-N 0.000 description 2
- 239000008049 TAE buffer Substances 0.000 description 2
- 108091036066 Three prime untranslated region Proteins 0.000 description 2
- 239000013504 Triton X-100 Substances 0.000 description 2
- 108090000631 Trypsin Proteins 0.000 description 2
- 102000004142 Trypsin Human genes 0.000 description 2
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 2
- 238000002835 absorbance Methods 0.000 description 2
- 238000010521 absorption reaction Methods 0.000 description 2
- HGEVZDLYZYVYHD-UHFFFAOYSA-N acetic acid;2-amino-2-(hydroxymethyl)propane-1,3-diol;2-[2-[bis(carboxymethyl)amino]ethyl-(carboxymethyl)amino]acetic acid Chemical compound CC(O)=O.OCC(N)(CO)CO.OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O HGEVZDLYZYVYHD-UHFFFAOYSA-N 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 125000000539 amino acid group Chemical group 0.000 description 2
- 235000012538 ammonium bicarbonate Nutrition 0.000 description 2
- 239000001099 ammonium carbonate Substances 0.000 description 2
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 2
- 229960000723 ampicillin Drugs 0.000 description 2
- 238000000137 annealing Methods 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- 125000000613 asparagine group Chemical group N[C@@H](CC(N)=O)C(=O)* 0.000 description 2
- 230000035578 autophosphorylation Effects 0.000 description 2
- 238000000376 autoradiography Methods 0.000 description 2
- 108010058966 bacteriophage T7 induced DNA polymerase Proteins 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 238000012742 biochemical analysis Methods 0.000 description 2
- 230000037396 body weight Effects 0.000 description 2
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 2
- 230000011712 cell development Effects 0.000 description 2
- 230000006727 cell loss Effects 0.000 description 2
- 230000003833 cell viability Effects 0.000 description 2
- 239000007795 chemical reaction product Substances 0.000 description 2
- 238000004140 cleaning Methods 0.000 description 2
- 238000003776 cleavage reaction Methods 0.000 description 2
- 238000004440 column chromatography Methods 0.000 description 2
- 238000012875 competitive assay Methods 0.000 description 2
- 239000012141 concentrate Substances 0.000 description 2
- 230000001143 conditioned effect Effects 0.000 description 2
- SUYVUBYJARFZHO-RRKCRQDMSA-N dATP Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@H]1C[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 SUYVUBYJARFZHO-RRKCRQDMSA-N 0.000 description 2
- SUYVUBYJARFZHO-UHFFFAOYSA-N dATP Natural products C1=NC=2C(N)=NC=NC=2N1C1CC(O)C(COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 SUYVUBYJARFZHO-UHFFFAOYSA-N 0.000 description 2
- 238000011033 desalting Methods 0.000 description 2
- MXHRCPNRJAMMIM-UHFFFAOYSA-N desoxyuridine Natural products C1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 MXHRCPNRJAMMIM-UHFFFAOYSA-N 0.000 description 2
- 229960002086 dextran Drugs 0.000 description 2
- 229960000633 dextran sulfate Drugs 0.000 description 2
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 2
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 2
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 2
- 210000000969 egg white Anatomy 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 229940116977 epidermal growth factor Drugs 0.000 description 2
- 239000013604 expression vector Substances 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 239000000835 fiber Substances 0.000 description 2
- 238000005194 fractionation Methods 0.000 description 2
- 239000011491 glass wool Substances 0.000 description 2
- RFDAIACWWDREDC-FRVQLJSFSA-N glycocholic acid Chemical compound C([C@H]1C[C@H]2O)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(=O)NCC(O)=O)C)[C@@]2(C)[C@@H](O)C1 RFDAIACWWDREDC-FRVQLJSFSA-N 0.000 description 2
- 210000005260 human cell Anatomy 0.000 description 2
- 230000002209 hydrophobic effect Effects 0.000 description 2
- 238000003384 imaging method Methods 0.000 description 2
- 238000007654 immersion Methods 0.000 description 2
- 102000018358 immunoglobulin Human genes 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- 239000003112 inhibitor Substances 0.000 description 2
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 239000012139 lysis buffer Substances 0.000 description 2
- 239000011159 matrix material Substances 0.000 description 2
- 238000005259 measurement Methods 0.000 description 2
- 238000002844 melting Methods 0.000 description 2
- 210000000274 microglia Anatomy 0.000 description 2
- 230000035772 mutation Effects 0.000 description 2
- 230000008764 nerve damage Effects 0.000 description 2
- 230000001537 neural effect Effects 0.000 description 2
- 230000004770 neurodegeneration Effects 0.000 description 2
- 208000015122 neurodegenerative disease Diseases 0.000 description 2
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 2
- 230000002093 peripheral effect Effects 0.000 description 2
- 210000001428 peripheral nervous system Anatomy 0.000 description 2
- 208000027232 peripheral nervous system disease Diseases 0.000 description 2
- 208000033808 peripheral neuropathy Diseases 0.000 description 2
- 229940080469 phosphocellulose Drugs 0.000 description 2
- 239000011574 phosphorus Substances 0.000 description 2
- 229910052698 phosphorus Inorganic materials 0.000 description 2
- DCWXELXMIBXGTH-UHFFFAOYSA-N phosphotyrosine Chemical compound OC(=O)C(N)CC1=CC=C(OP(O)(O)=O)C=C1 DCWXELXMIBXGTH-UHFFFAOYSA-N 0.000 description 2
- 239000013600 plasmid vector Substances 0.000 description 2
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 2
- -1 polyoxy Polymers 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 238000004321 preservation Methods 0.000 description 2
- 238000011002 quantification Methods 0.000 description 2
- 238000004445 quantitative analysis Methods 0.000 description 2
- 238000005215 recombination Methods 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 238000004366 reverse phase liquid chromatography Methods 0.000 description 2
- 239000012723 sample buffer Substances 0.000 description 2
- 239000012047 saturated solution Substances 0.000 description 2
- 230000007017 scission Effects 0.000 description 2
- 230000001953 sensory effect Effects 0.000 description 2
- 239000001632 sodium acetate Substances 0.000 description 2
- 235000017281 sodium acetate Nutrition 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- 238000001228 spectrum Methods 0.000 description 2
- 238000003756 stirring Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 239000000725 suspension Substances 0.000 description 2
- 208000024891 symptom Diseases 0.000 description 2
- 230000002194 synthesizing effect Effects 0.000 description 2
- 238000010257 thawing Methods 0.000 description 2
- 238000002560 therapeutic procedure Methods 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 102000035160 transmembrane proteins Human genes 0.000 description 2
- 108091005703 transmembrane proteins Proteins 0.000 description 2
- LWIHDJKSTIGBAC-UHFFFAOYSA-K tripotassium phosphate Chemical compound [K+].[K+].[K+].[O-]P([O-])([O-])=O LWIHDJKSTIGBAC-UHFFFAOYSA-K 0.000 description 2
- 239000012588 trypsin Substances 0.000 description 2
- 210000004881 tumor cell Anatomy 0.000 description 2
- 241001515965 unidentified phage Species 0.000 description 2
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 2
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 1
- PQMRRAQXKWFYQN-UHFFFAOYSA-N 1-phenyl-2-sulfanylideneimidazolidin-4-one Chemical compound S=C1NC(=O)CN1C1=CC=CC=C1 PQMRRAQXKWFYQN-UHFFFAOYSA-N 0.000 description 1
- GZCWLCBFPRFLKL-UHFFFAOYSA-N 1-prop-2-ynoxypropan-2-ol Chemical compound CC(O)COCC#C GZCWLCBFPRFLKL-UHFFFAOYSA-N 0.000 description 1
- GOJUJUVQIVIZAV-UHFFFAOYSA-N 2-amino-4,6-dichloropyrimidine-5-carbaldehyde Chemical group NC1=NC(Cl)=C(C=O)C(Cl)=N1 GOJUJUVQIVIZAV-UHFFFAOYSA-N 0.000 description 1
- VADKRMSMGWJZCF-UHFFFAOYSA-N 2-bromophenol Chemical compound OC1=CC=CC=C1Br VADKRMSMGWJZCF-UHFFFAOYSA-N 0.000 description 1
- 102100027324 2-hydroxyacyl-CoA lyase 1 Human genes 0.000 description 1
- CIHKVMHPDDJIIP-UHFFFAOYSA-N 2-methylperoxybenzoic acid Chemical compound COOC1=CC=CC=C1C(O)=O CIHKVMHPDDJIIP-UHFFFAOYSA-N 0.000 description 1
- NKDFYOWSKOHCCO-YPVLXUMRSA-N 20-hydroxyecdysone Chemical compound C1[C@@H](O)[C@@H](O)C[C@]2(C)[C@@H](CC[C@@]3([C@@H]([C@@](C)(O)[C@H](O)CCC(C)(O)C)CC[C@]33O)C)C3=CC(=O)[C@@H]21 NKDFYOWSKOHCCO-YPVLXUMRSA-N 0.000 description 1
- XZKIHKMTEMTJQX-UHFFFAOYSA-N 4-Nitrophenyl Phosphate Chemical compound OP(O)(=O)OC1=CC=C([N+]([O-])=O)C=C1 XZKIHKMTEMTJQX-UHFFFAOYSA-N 0.000 description 1
- 102000013563 Acid Phosphatase Human genes 0.000 description 1
- 108010051457 Acid Phosphatase Proteins 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- VHUUQVKOLVNVRT-UHFFFAOYSA-N Ammonium hydroxide Chemical compound [NH4+].[OH-] VHUUQVKOLVNVRT-UHFFFAOYSA-N 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- BTBUEUYNUDRHOZ-UHFFFAOYSA-N Borate Chemical compound [O-]B([O-])[O-] BTBUEUYNUDRHOZ-UHFFFAOYSA-N 0.000 description 1
- 101100499406 Bos taurus DNA2 gene Proteins 0.000 description 1
- 101100120045 Bos taurus FGF1 gene Proteins 0.000 description 1
- 238000010599 BrdU assay Methods 0.000 description 1
- CPELXLSAUQHCOX-UHFFFAOYSA-M Bromide Chemical compound [Br-] CPELXLSAUQHCOX-UHFFFAOYSA-M 0.000 description 1
- 238000006037 Brook Silaketone rearrangement reaction Methods 0.000 description 1
- 108091016585 CD44 antigen Proteins 0.000 description 1
- 238000002238 CM-cellulose chromatography Methods 0.000 description 1
- 244000025254 Cannabis sativa Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 102100031196 Choriogonadotropin subunit beta 3 Human genes 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- KDXKERNSBIXSRK-RXMQYKEDSA-N D-lysine Chemical compound NCCCC[C@@H](N)C(O)=O KDXKERNSBIXSRK-RXMQYKEDSA-N 0.000 description 1
- 101150074155 DHFR gene Proteins 0.000 description 1
- 230000004544 DNA amplification Effects 0.000 description 1
- 108010008286 DNA nucleotidylexotransferase Proteins 0.000 description 1
- 238000012270 DNA recombination Methods 0.000 description 1
- 102100029764 DNA-directed DNA/RNA polymerase mu Human genes 0.000 description 1
- 108010054576 Deoxyribonuclease EcoRI Proteins 0.000 description 1
- 102100024746 Dihydrofolate reductase Human genes 0.000 description 1
- 108090000204 Dipeptidase 1 Proteins 0.000 description 1
- 102000001301 EGF receptor Human genes 0.000 description 1
- 108060006698 EGF receptor Proteins 0.000 description 1
- 241000701832 Enterobacteria phage T3 Species 0.000 description 1
- 208000034454 F12-related hereditary angioedema with normal C1Inh Diseases 0.000 description 1
- 102100031706 Fibroblast growth factor 1 Human genes 0.000 description 1
- 108090000379 Fibroblast growth factor 2 Proteins 0.000 description 1
- 102100024785 Fibroblast growth factor 2 Human genes 0.000 description 1
- 241000287828 Gallus gallus Species 0.000 description 1
- 102100032864 General transcription factor IIH subunit 2 Human genes 0.000 description 1
- 108010051815 Glutamyl endopeptidase Proteins 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 239000000579 Gonadotropin-Releasing Hormone Substances 0.000 description 1
- 108060003393 Granulin Proteins 0.000 description 1
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 description 1
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 description 1
- 102000008055 Heparan Sulfate Proteoglycans Human genes 0.000 description 1
- 229920002971 Heparan sulfate Polymers 0.000 description 1
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 1
- 229920000209 Hexadimethrine bromide Polymers 0.000 description 1
- 102100037907 High mobility group protein B1 Human genes 0.000 description 1
- 101710168537 High mobility group protein B1 Proteins 0.000 description 1
- 102100022128 High mobility group protein B2 Human genes 0.000 description 1
- 101710168540 High mobility group protein B2 Proteins 0.000 description 1
- 241000282412 Homo Species 0.000 description 1
- 101001009252 Homo sapiens 2-hydroxyacyl-CoA lyase 1 Proteins 0.000 description 1
- 101000776619 Homo sapiens Choriogonadotropin subunit beta 3 Proteins 0.000 description 1
- 101000599940 Homo sapiens Interferon gamma Proteins 0.000 description 1
- 101000582546 Homo sapiens Methylosome protein 50 Proteins 0.000 description 1
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 101000740205 Homo sapiens Sal-like protein 1 Proteins 0.000 description 1
- 241000714260 Human T-lymphotropic virus 1 Species 0.000 description 1
- 102000004877 Insulin Human genes 0.000 description 1
- 108090001061 Insulin Proteins 0.000 description 1
- 108010050904 Interferons Proteins 0.000 description 1
- 102000014150 Interferons Human genes 0.000 description 1
- 108090001005 Interleukin-6 Proteins 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 229930182816 L-glutamine Natural products 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 1
- 108090000364 Ligases Proteins 0.000 description 1
- 102000003960 Ligases Human genes 0.000 description 1
- 108010073521 Luteinizing Hormone Proteins 0.000 description 1
- 102000009151 Luteinizing Hormone Human genes 0.000 description 1
- 229910021380 Manganese Chloride Inorganic materials 0.000 description 1
- GLFNIEUTAYBVOC-UHFFFAOYSA-L Manganese chloride Chemical compound Cl[Mn]Cl GLFNIEUTAYBVOC-UHFFFAOYSA-L 0.000 description 1
- 108091027974 Mature messenger RNA Proteins 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- OKIZCWYLBDKLSU-UHFFFAOYSA-M N,N,N-Trimethylmethanaminium chloride Chemical compound [Cl-].C[N+](C)(C)C OKIZCWYLBDKLSU-UHFFFAOYSA-M 0.000 description 1
- CHJJGSNFBQVOTG-UHFFFAOYSA-N N-methyl-guanidine Natural products CNC(N)=N CHJJGSNFBQVOTG-UHFFFAOYSA-N 0.000 description 1
- 229920002274 Nalgene Polymers 0.000 description 1
- 201000004404 Neurofibroma Diseases 0.000 description 1
- 208000009905 Neurofibromatoses Diseases 0.000 description 1
- 102000011931 Nucleoproteins Human genes 0.000 description 1
- 108010061100 Nucleoproteins Proteins 0.000 description 1
- 108700026244 Open Reading Frames Proteins 0.000 description 1
- 108010058846 Ovalbumin Proteins 0.000 description 1
- 208000012868 Overgrowth Diseases 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 108700020978 Proto-Oncogene Proteins 0.000 description 1
- 102000052575 Proto-Oncogene Human genes 0.000 description 1
- 108091034057 RNA (poly(A)) Proteins 0.000 description 1
- 239000012979 RPMI medium Substances 0.000 description 1
- 101001010820 Rattus norvegicus Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 108010039491 Ricin Proteins 0.000 description 1
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 1
- 239000012722 SDS sample buffer Substances 0.000 description 1
- 102100037204 Sal-like protein 1 Human genes 0.000 description 1
- 101000857870 Squalus acanthias Gonadoliberin Proteins 0.000 description 1
- 239000012505 Superdex™ Substances 0.000 description 1
- 108090000054 Syndecan-2 Proteins 0.000 description 1
- YZCKVEUIGOORGS-NJFSPNSNSA-N Tritium Chemical compound [3H] YZCKVEUIGOORGS-NJFSPNSNSA-N 0.000 description 1
- 108091023045 Untranslated Region Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- 206010052428 Wound Diseases 0.000 description 1
- ZKHQWZAMYRWXGA-KNYAHOBESA-N [[(2r,3s,4r,5r)-5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl] dihydroxyphosphoryl hydrogen phosphate Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](COP(O)(=O)OP(O)(=O)O[32P](O)(O)=O)[C@@H](O)[C@H]1O ZKHQWZAMYRWXGA-KNYAHOBESA-N 0.000 description 1
- 230000002159 abnormal effect Effects 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 239000011149 active material Substances 0.000 description 1
- 201000003352 adrenal gland pheochromocytoma Diseases 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 239000000443 aerosol Substances 0.000 description 1
- 239000000556 agonist Substances 0.000 description 1
- 238000003277 amino acid sequence analysis Methods 0.000 description 1
- 235000019270 ammonium chloride Nutrition 0.000 description 1
- 239000000908 ammonium hydroxide Substances 0.000 description 1
- 239000012491 analyte Substances 0.000 description 1
- 238000012442 analytical experiment Methods 0.000 description 1
- 210000004102 animal cell Anatomy 0.000 description 1
- 238000005349 anion exchange Methods 0.000 description 1
- 238000005571 anion exchange chromatography Methods 0.000 description 1
- 239000005557 antagonist Substances 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 239000000427 antigen Substances 0.000 description 1
- 108091007433 antigens Proteins 0.000 description 1
- 102000036639 antigens Human genes 0.000 description 1
- 239000008346 aqueous phase Substances 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 210000002469 basement membrane Anatomy 0.000 description 1
- 108010066058 beta Subunit Luteinizing Hormone Proteins 0.000 description 1
- 230000002457 bidirectional effect Effects 0.000 description 1
- 230000005540 biological transmission Effects 0.000 description 1
- KGBXLFKZBHKPEV-UHFFFAOYSA-N boric acid Chemical compound OB(O)O KGBXLFKZBHKPEV-UHFFFAOYSA-N 0.000 description 1
- 239000004327 boric acid Substances 0.000 description 1
- 210000000133 brain stem Anatomy 0.000 description 1
- 239000007853 buffer solution Substances 0.000 description 1
- 244000309464 bull Species 0.000 description 1
- AIYUHDOJVYHVIT-UHFFFAOYSA-M caesium chloride Inorganic materials [Cl-].[Cs+] AIYUHDOJVYHVIT-UHFFFAOYSA-M 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 244000309466 calf Species 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 239000002775 capsule Substances 0.000 description 1
- 239000004202 carbamide Substances 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 230000009920 chelation Effects 0.000 description 1
- 238000011210 chromatographic step Methods 0.000 description 1
- 239000012504 chromatography matrix Substances 0.000 description 1
- 238000011097 chromatography purification Methods 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 238000005352 clarification Methods 0.000 description 1
- 238000012411 cloning technique Methods 0.000 description 1
- 238000000975 co-precipitation Methods 0.000 description 1
- 230000002860 competitive effect Effects 0.000 description 1
- 230000006957 competitive inhibition Effects 0.000 description 1
- 230000000295 complement effect Effects 0.000 description 1
- 239000002131 composite material Substances 0.000 description 1
- 230000003750 conditioning effect Effects 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 238000004132 cross linking Methods 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- ATDGTVJJHBUTRL-UHFFFAOYSA-N cyanogen bromide Chemical compound BrC#N ATDGTVJJHBUTRL-UHFFFAOYSA-N 0.000 description 1
- 150000001945 cysteines Chemical class 0.000 description 1
- LEVWYRKDKASIDU-IMJSIDKUSA-N cystine group Chemical group C([C@@H](C(=O)O)N)SSC[C@@H](C(=O)O)N LEVWYRKDKASIDU-IMJSIDKUSA-N 0.000 description 1
- 238000007405 data analysis Methods 0.000 description 1
- 239000007857 degradation product Substances 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 238000001739 density measurement Methods 0.000 description 1
- 229940009976 deoxycholate Drugs 0.000 description 1
- KXGVEGMKQFWNSR-LLQZFEROSA-N deoxycholic acid Chemical compound C([C@H]1CC2)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(O)=O)C)[C@@]2(C)[C@@H](O)C1 KXGVEGMKQFWNSR-LLQZFEROSA-N 0.000 description 1
- 230000008021 deposition Effects 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- 206010012601 diabetes mellitus Diseases 0.000 description 1
- 238000000502 dialysis Methods 0.000 description 1
- 239000005546 dideoxynucleotide Substances 0.000 description 1
- 108020001096 dihydrofolate reductase Proteins 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- SWSQBOPZIKWTGO-UHFFFAOYSA-N dimethylaminoamidine Natural products CN(C)C(N)=N SWSQBOPZIKWTGO-UHFFFAOYSA-N 0.000 description 1
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 1
- 230000008034 disappearance Effects 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037765 diseases and disorders Diseases 0.000 description 1
- 208000035475 disorder Diseases 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 239000012149 elution buffer Substances 0.000 description 1
- 230000013020 embryo development Effects 0.000 description 1
- 239000006167 equilibration buffer Substances 0.000 description 1
- 150000002148 esters Chemical class 0.000 description 1
- 238000012869 ethanol precipitation Methods 0.000 description 1
- ZMMJGEGLRURXTF-UHFFFAOYSA-N ethidium bromide Chemical compound [Br-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CC)=C1C1=CC=CC=C1 ZMMJGEGLRURXTF-UHFFFAOYSA-N 0.000 description 1
- 229960005542 ethidium bromide Drugs 0.000 description 1
- DEFVIWRASFVYLL-UHFFFAOYSA-N ethylene glycol bis(2-aminoethyl)tetraacetic acid Chemical compound OC(=O)CN(CC(O)=O)CCOCCOCCN(CC(O)=O)CC(O)=O DEFVIWRASFVYLL-UHFFFAOYSA-N 0.000 description 1
- 239000005038 ethylene vinyl acetate Substances 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 206010016165 failure to thrive Diseases 0.000 description 1
- 238000011049 filling Methods 0.000 description 1
- 239000000706 filtrate Substances 0.000 description 1
- 210000001652 frontal lobe Anatomy 0.000 description 1
- 230000006870 function Effects 0.000 description 1
- 238000001641 gel filtration chromatography Methods 0.000 description 1
- 238000012239 gene modification Methods 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 230000005017 genetic modification Effects 0.000 description 1
- 235000013617 genetically modified food Nutrition 0.000 description 1
- 239000011521 glass Substances 0.000 description 1
- XLXSAKCOAKORKW-AQJXLSMYSA-N gonadorelin Chemical compound C([C@@H](C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N1[C@@H](CCC1)C(=O)NCC(N)=O)NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H]1NC(=O)CC1)C1=CC=C(O)C=C1 XLXSAKCOAKORKW-AQJXLSMYSA-N 0.000 description 1
- 229940035638 gonadotropin-releasing hormone Drugs 0.000 description 1
- 230000005484 gravity Effects 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 208000016861 hereditary angioedema type 3 Diseases 0.000 description 1
- 108010044853 histidine-rich proteins Proteins 0.000 description 1
- 210000003016 hypothalamus Anatomy 0.000 description 1
- 239000005457 ice water Substances 0.000 description 1
- 230000002163 immunogen Effects 0.000 description 1
- 229940027941 immunoglobulin g Drugs 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 230000002779 inactivation Effects 0.000 description 1
- 239000000411 inducer Substances 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 239000002198 insoluble material Substances 0.000 description 1
- 229940125396 insulin Drugs 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- 229940079322 interferon Drugs 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 238000007917 intracranial administration Methods 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000007913 intrathecal administration Methods 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 238000007914 intraventricular administration Methods 0.000 description 1
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 1
- 210000003292 kidney cell Anatomy 0.000 description 1
- 238000011005 laboratory method Methods 0.000 description 1
- JJTUDXZGHPGLLC-UHFFFAOYSA-N lactide Chemical compound CC1OC(=O)C(C)OC1=O JJTUDXZGHPGLLC-UHFFFAOYSA-N 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 239000010410 layer Substances 0.000 description 1
- 230000031700 light absorption Effects 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 239000007788 liquid Substances 0.000 description 1
- 239000012160 loading buffer Substances 0.000 description 1
- 238000011068 loading method Methods 0.000 description 1
- 238000000464 low-speed centrifugation Methods 0.000 description 1
- 229940040129 luteinizing hormone Drugs 0.000 description 1
- 210000002540 macrophage Anatomy 0.000 description 1
- 229910001629 magnesium chloride Inorganic materials 0.000 description 1
- 238000012423 maintenance Methods 0.000 description 1
- 230000036210 malignancy Effects 0.000 description 1
- 239000011565 manganese chloride Substances 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- 239000003607 modifier Substances 0.000 description 1
- 230000004660 morphological change Effects 0.000 description 1
- 201000006417 multiple sclerosis Diseases 0.000 description 1
- MQUQNUAYKLCRME-UHFFFAOYSA-N n-(4-chloro-3-oxo-1-phenylbutan-2-yl)-4-methylbenzenesulfonamide Chemical compound C1=CC(C)=CC=C1S(=O)(=O)NC(C(=O)CCl)CC1=CC=CC=C1 MQUQNUAYKLCRME-UHFFFAOYSA-N 0.000 description 1
- 150000002790 naphthalenes Chemical class 0.000 description 1
- 239000007923 nasal drop Substances 0.000 description 1
- 229940100662 nasal drops Drugs 0.000 description 1
- 208000023833 nerve sheath neoplasm Diseases 0.000 description 1
- 201000004931 neurofibromatosis Diseases 0.000 description 1
- 230000007971 neurological deficit Effects 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- JPXMTWWFLBLUCD-UHFFFAOYSA-N nitro blue tetrazolium(2+) Chemical compound COC1=CC(C=2C=C(OC)C(=CC=2)[N+]=2N(N=C(N=2)C=2C=CC=CC=2)C=2C=CC(=CC=2)[N+]([O-])=O)=CC=C1[N+]1=NC(C=2C=CC=CC=2)=NN1C1=CC=C([N+]([O-])=O)C=C1 JPXMTWWFLBLUCD-UHFFFAOYSA-N 0.000 description 1
- 108091027963 non-coding RNA Proteins 0.000 description 1
- 102000042567 non-coding RNA Human genes 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 102000027450 oncoproteins Human genes 0.000 description 1
- 108091008819 oncoproteins Proteins 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 230000003204 osmotic effect Effects 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 230000001575 pathological effect Effects 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 230000001991 pathophysiological effect Effects 0.000 description 1
- 210000003024 peritoneal macrophage Anatomy 0.000 description 1
- 239000000825 pharmaceutical preparation Substances 0.000 description 1
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 230000000704 physical effect Effects 0.000 description 1
- 210000003635 pituitary gland Anatomy 0.000 description 1
- 229920003023 plastic Polymers 0.000 description 1
- 239000004033 plastic Substances 0.000 description 1
- 239000002574 poison Substances 0.000 description 1
- 231100000614 poison Toxicity 0.000 description 1
- 229920001200 poly(ethylene-vinyl acetate) Polymers 0.000 description 1
- 229920001515 polyalkylene glycol Polymers 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 229920001451 polypropylene glycol Polymers 0.000 description 1
- 229920006316 polyvinylpyrrolidine Polymers 0.000 description 1
- 229910000160 potassium phosphate Inorganic materials 0.000 description 1
- 235000011009 potassium phosphates Nutrition 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 239000002244 precipitate Substances 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 230000002265 prevention Effects 0.000 description 1
- 230000003449 preventive effect Effects 0.000 description 1
- 230000037452 priming Effects 0.000 description 1
- 230000009696 proliferative response Effects 0.000 description 1
- 235000019833 protease Nutrition 0.000 description 1
- 230000009822 protein phosphorylation Effects 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 239000013014 purified material Substances 0.000 description 1
- 239000012264 purified product Substances 0.000 description 1
- 239000002994 raw material Substances 0.000 description 1
- 239000000376 reactant Substances 0.000 description 1
- 230000009257 reactivity Effects 0.000 description 1
- 108091008598 receptor tyrosine kinases Proteins 0.000 description 1
- 102000027426 receptor tyrosine kinases Human genes 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 230000001105 regulatory effect Effects 0.000 description 1
- 238000007634 remodeling Methods 0.000 description 1
- 230000008458 response to injury Effects 0.000 description 1
- 210000001995 reticulocyte Anatomy 0.000 description 1
- 238000010839 reverse transcription Methods 0.000 description 1
- 230000003248 secreting effect Effects 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 210000002265 sensory receptor cell Anatomy 0.000 description 1
- 102000027509 sensory receptors Human genes 0.000 description 1
- 108091008691 sensory receptors Proteins 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- CMZUMMUJMWNLFH-UHFFFAOYSA-N sodium metavanadate Chemical compound [Na+].[O-][V](=O)=O CMZUMMUJMWNLFH-UHFFFAOYSA-N 0.000 description 1
- 239000012064 sodium phosphate buffer Substances 0.000 description 1
- 238000001179 sorption measurement Methods 0.000 description 1
- 238000010183 spectrum analysis Methods 0.000 description 1
- 210000000278 spinal cord Anatomy 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 101150075348 sta gene Proteins 0.000 description 1
- 239000008223 sterile water Substances 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 208000011580 syndromic disease Diseases 0.000 description 1
- 239000012085 test solution Substances 0.000 description 1
- 229940124597 therapeutic agent Drugs 0.000 description 1
- 230000000451 tissue damage Effects 0.000 description 1
- 231100000827 tissue damage Toxicity 0.000 description 1
- 230000017423 tissue regeneration Effects 0.000 description 1
- 229950003937 tolonium Drugs 0.000 description 1
- HNONEKILPDHFOL-UHFFFAOYSA-M tolonium chloride Chemical compound [Cl-].C1=C(C)C(N)=CC2=[S+]C3=CC(N(C)C)=CC=C3N=C21 HNONEKILPDHFOL-UHFFFAOYSA-M 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- PIEPQKCYPFFYMG-UHFFFAOYSA-N tris acetate Chemical compound CC(O)=O.OCC(N)(CO)CO PIEPQKCYPFFYMG-UHFFFAOYSA-N 0.000 description 1
- 239000003656 tris buffered saline Substances 0.000 description 1
- 229910052722 tritium Inorganic materials 0.000 description 1
- GPRLSGONYQIRFK-MNYXATJNSA-N triton Chemical compound [3H+] GPRLSGONYQIRFK-MNYXATJNSA-N 0.000 description 1
- 235000015112 vegetable and seed oil Nutrition 0.000 description 1
- 239000008158 vegetable oil Substances 0.000 description 1
- 210000005172 vertebrate brain Anatomy 0.000 description 1
- 239000011534 wash buffer Substances 0.000 description 1
- 230000029663 wound healing Effects 0.000 description 1
- 229910000166 zirconium phosphate Inorganic materials 0.000 description 1
Images

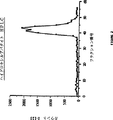


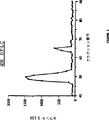


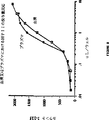






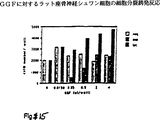


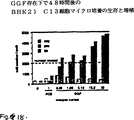
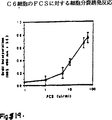






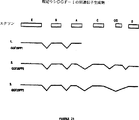







































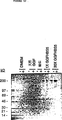

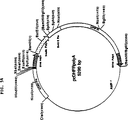
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4702—Regulators; Modulating activity
- C07K14/4705—Regulators; Modulating activity stimulating, promoting or activating activity
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/475—Growth factors; Growth regulators
- C07K14/4756—Neuregulins, i.e. p185erbB2 ligands, glial growth factor, heregulin, ARIA, neu differentiation factor
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/82—Translation products from oncogenes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/22—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against growth factors ; against growth regulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2799/00—Uses of viruses
- C12N2799/02—Uses of viruses as vector
- C12N2799/021—Uses of viruses as vector for the expression of a heterologous nucleic acid
- C12N2799/026—Uses of viruses as vector for the expression of a heterologous nucleic acid where the vector is derived from a baculovirus
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- General Chemical & Material Sciences (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Toxicology (AREA)
- Zoology (AREA)
- Pharmacology & Pharmacy (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Engineering & Computer Science (AREA)
- Oncology (AREA)
- Immunology (AREA)
- Neurology (AREA)
- Biomedical Technology (AREA)
- Dermatology (AREA)
- Neurosurgery (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Saccharide Compounds (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Acyclic And Carbocyclic Compounds In Medicinal Compositions (AREA)
- Medicinal Preparation (AREA)
Description
関連出願の相互参照
本出願は、19本出願は、1992年10月23日出願の出願番号第07/965,173号と、1992年9月3日出願の出願番号第07/940,389号と、1992年6月30日出願の出願番号第07/907,138号と、1992年4月3日出願の出願番号第07/863,703号との一部継続出願である。
【0002】
発明の背景
本発明は、脊椎類に見られ、シュワン細胞を含むグリア細胞の細胞分裂誘発成長因子であるポリペプチドに関する。本発明は、更に、そのような因子を生成可能な方法と、そのような因子の治療的利用とにも関する。
【0003】
脊椎類のグリア細胞は、中枢及び末梢神経システムの特殊化した結合組織を構成している。重要なグリア細胞としてシュワン細胞があるが、これは、ニューロンの為の代謝支持体を提供するとともに、特定の末梢神経の神経軸索の周囲に髄鞘(ミエリン鞘)を提供することによって、個別の神経繊維を形成する。シュワン細胞は、ニューロンを支持し、近接する神経軸索の周囲において近接同心の膜層を形成し、その成長につれて前記軸索の周囲において捻れながら形成されることによってミエリン鞘効果を提供するものである。これらのミエリン鞘は、多くの神経線維の弱い部分であり、これらシュワン細胞の損傷、発育及び成長の不全が、数多くの末梢神経システムの疾患と障害を特徴付ける大幅な脱髄や神経退化に関連している可能性がある。神経システムの発達において、その細胞がその分裂と成長を制御するのに様々な因子を必要とすることが明らかになった。そして、近年、このような種々の因子が同定され、これらの内のいくつかはシュワン細胞の分裂と発達に影響することが判っている。
【0004】
従って、ブロックス(Brockes)他,interalia,in Neuroscience,4(1984)75−83は、ウシの脳と下垂体組織とからの抽出物質に存在するタンパク質成長因子について記載しており、これはグリア細胞成長因子(GGF)と命名された。この因子は、ウシ胎仔血清を10%含有するバックグラウンド培地において、培養ラットのシュワン細胞を刺激してこれを分裂させた。又、この因子は分子量が31,000ダルトンであり、容易にダイマー化することも記載されている。In Meth.Enz.,147(1987),217−225において、ブロックス(Brockes)は、GGFの、シュワン細胞に基づく分析について記載している。
【0005】
ブロックス(Brockes)他,上記、は、更に、GGFの外見上の均質状態への精製方法についても記載している。要約すると、ここに記載された一つの大規模精製方法は、凍結乾燥したウシの前頭葉の抽出と、それによって得られた物質をCMセルロースからのNaClグラジエント溶出を使用したクロマトグラフィーに関するものである。次に、ゲル濾過を、先ずウルトロゲル(Ultrogel)カラムで、次に、ホスホセルロースカラムで、そして最後に、小規模SDSゲル電気泳動法で行う。あるいは、前記CMセルロース物質を、ホスホセルロースカラムに直接に適用し、該カラムからのフラクションをプールして、調製天然ゲル電気泳動によって精製し、その後、最終的なSDSゲル電気泳動にかける。
【0006】
ブロックス(Brockes)他は、以前に報告されたゲル濾過実験(ブロックス(Brockes)他,J.Biol.Chem.255(1980)8374−8377)において、成長因子活性の大きなピークが、分子量56,000ダルトンで移動(migrate)することが観察され、これに対して、上述の手法の最初のものにおいては、分子量31,000においておもに活性が観察された、と述べている。GGFダイマーは、この実験において、主として、CMセルロースからのグラジエント溶出の生成物として除去されることが報告されている。
【0007】
ベンヴェニスト(Benveniste)他(PNAS,82(1885),3930−3934)は、Tリンパ球由来のグリア細胞発育誘発因子について記載している。この因子は、還元状態において、SDSゲルの見かけ上の分子量の変化を示す。
【0008】
キムラ(Kimura)他(Nature,348(1990),257−260)は、自ら神経線維腫由来成長因子(SDGF)と呼称する座骨神経鞘腫瘍から得られる因子について記載している。これらの著者は、反対に部分的に精製された下垂体フラクション含有のGGFが活性な状態においては、SDGFは、トリチウム標識化TdRのシュワン細胞への結合を刺激しないと述べている。SDGFの見かけ上の分子量は31,000ないし35,000である。
【0009】
デイヴィス(Davis)とストローバント(Stroobant)(J.Cell.Biol.,110(1990),1353〜1360)とは、多数の候補細胞分裂誘発物質について記載している。ラットのシュワン細胞を使用し、選択された候補物質が、FCS(ウシ胎仔血清)が10%存在して、フォルスコリン(forskolin)が存在する場合と、存在しない場合とにおいて、シュワン細胞へのDNA合成を刺激する能力をテストした。テストした因子の一つは、GGF−カルボキシメチルセルロースフラクション(GGF−CM)であり、これはFCSが存在する場合、フォルスコリン(forskolin)が存在及び不在の両方の場合において細胞分裂誘発作用を有していた。この研究に依り、フォルスコリンの存在下において、特に血小板由来成長因子(PDGF)が、シュワン細胞に対する潜在的細胞分裂誘発物質であることが判った。PDGFは、以前には、シュワン細胞に影響しないと考えられていたものである。
【0010】
ホルムズ(Holmes)他,Science(1992)256:1205及びウェン(Wen)他,Cell(1992)69:559は、レセプター(p185erbB2)に結合するタンパク質をコード化するDNA配列がいくつかのヒトの腫瘍に関連していることを示している。
【0011】
前記p185erbB2タンパク質は、チロシンキナーゼ活性を備えた185キロダルトン膜長のタンパク質である。そしてこのタンパク質は、erbB2プロト−がん遺伝子によってコード化される(ヤーデン(Yarden)及びユルリッチ(Ullrich)Ann.Rev.Biochem.57:433(1988))。前記rbB2遺伝子は、HER−2(ヒト細胞)、あるいはeneu(ラット細胞)とも呼称されるが、上皮細胞成長因子(EGF)のレセプターに密接に関連している。最近の証拠に依れば、p185erbB2と作用する(そしてそのキナーゼを活性化する)タンパク質は、p185erbB2を有する細胞の増殖を誘発することが示されている(ホルムズ(Holmes)他,Science 256:1205(1992);ドバシ他,Proc.Natl.Acad.Sci.88:8582(1991);ルプ(Lupu)他,Proc.Natl.Acad.Sci.89:2287(1992))。更に、p185erbB2結合タンパク質をコード化する前記遺伝子が、それぞれ長さが異なり、かつ、いくつかの共通のペプチド配列とユニークなペプチド配列とを含む一連のタンパク質を生成する多くの、大きさとスプライシングとが異なったRNA転写体を生成することが明らかである。これは、ヒトの乳ガン(MDA−MB−231)から得られるスプライシングが異なったRNA転写体によって証明されている(ホルムズ(Holmes)他,Science 256:1205(1992))。更に、これは(ここに記載されているように)p185erbB2レセプターのためのリガンドとして作用する広範囲のタンパク質によっても証明されている(下記参照)。
【0012】
発明の要旨
一般に、本発明は、グリア細胞(特に、シュワン細胞と中枢神経システムのグリア)の分裂を刺激する方法と、更に、そのようなグリア細胞の分裂誘発活性を示す新規なタンパク質とに関する。更に、これらのタンパク質とこれらタンパク質と、関連タンパク質とに結合する抗体をコード化するDNAも提供される。
【0013】
本発明の新規なタンパク質は、公知のポリペプチドをコード化する配列の別のスプライス生成物を含む。一般に、これらの公知タンパク質は、前記GGF/p185erbB2族タンパク質のメンバーである。
【0014】
より詳しくは、本発明は、所定の式のポリペプチドと、これらのポリペプチドをコード化するDNA配列とを提供する。即ち、これらポリペプチドは、以下の式を有する。
【0015】
WYBAZCX
ここで、WYBAZCXは、図31に示されるアミノ酸配列から成り(配列認識番号136〜139、141〜147、160、161)、Wは、ポリペプチドセグメントFを有するかあるいは有さず、Yは、ポリペプチドセグメントEを有するかあるいは有さず、Zは、ポリペプチドセグメントGを有するかあるいは有さず、そして、ここにXはポリペプチドセグメントC/D HKL,C/D H,C/D HL,C/D D,C/D’HL,C/D’HKL,C/D’H,C/D’D,C/D C/D’HKL,C/D C/D’H,C/D C/D’HL,C/D C/D’D,C/D D’H,C/D D’HL,C/D D’HKL,C/D’D’H,C/D’D’HL,C/D’D’HKL,C/D C/D’D’H,C/D C/D’D’HL,またはC/D C/D’D’HKLを含み、
以下のいずれかの条件を満たす、
a)F、Y、B、A、Z、C又はXの少なくとも一つは、ウシ由来である、
b)YはポリペプチドセグメントEであるか、又は、
c)XはポリペプチドセグメントC/D HKL,C/D D,C/D’HKL,C/D C/D’HKL,C/D C/D’D,C/D D’H,C/D D’HL,C/D D’HKL,C/D’D’H,C/D’D’HKL,C/D C/D’D’H,C/D C/D’D’HL,C/D C/D’HKL,C/D’H,C/D C/D’H,またはC/D C/D’HLを含む。
【0016】
更に、本発明は、コード化セグメント5’FBA3’と、図31に示されるアミノ酸配列(配列認識番号136、138、139)を有する対応のポリペプチドセグメントを備えたDNA配列も含む。
【0017】
前記DNA配列は、コード化セグメント5’FBA3’と、図31に示されるアミノ酸配列(配列認識番号136,138,140)を有する対応のポリペプチドセグメントを含む。
【0018】
前記DNA配列は、コード化セグメント5’FEBA3’と、図31に示されるアミノ酸配列(配列認識番号136〜139)を有する対応のポリペプチドセグメントを含む。
【0019】
前記DNA配列は、コード化セグメント5’FEBA3’と、図31に示されるアミノ酸配列(配列認識番号136〜138、140)を有する対応のポリペプチドセグメントを含む。そして、前記DNA配列は、CGF2HBS5 cDNAクローンのポリペプチドコード化セグメントを含む(ATCC Deposit No.75298,deposited1992年9月2日)。
【0020】
本発明は、更に、式FBA,FEBA,FBA’,FEBA’のペプチドと、これらペプチドをコード化するDNA配列とを含み、前記ポリペプチドセグメントは、図31において、それぞれ、配列認識番号(136,138及び139)、(136〜139)と(136、138及び140)、及び(136〜138及び140)とで示されるアミノ酸配列に対応する。精製GGF−IIポリペプチド(配列認識番号170)である前記ポリペプチドも本発明の一部に含まれる。
【0021】
更に、本発明の一態様は、中枢神経システムのグリア、特に乏突起膠細胞、小グリア細胞及び星状膠細胞、の治療に有用なペプチドと、これらのペプチドをコード化するDNAと、更にこれらのペプチドの投与方法を含む。
【0022】
本発明は、更に、前記定義のアミノ酸配列をコード化するDNA配列を含むベクターを含む。又、前述のアミノ酸配列をコード化する単離DNAを含有するホスト細胞も含まれる。本発明は、更に、前記p185erbB2レセプターを結合し、生体内及び/又は生体外でグリア細胞分裂を刺激する化合物を含む。
【0023】
また、前述の新規なペプチドに対する抗体も本発明の一部として含まれる。更に、ここに記載のすべてのペプチドに対する抗体は、ここに記載のポリペプチドの精製に利用可能である。又、これらのポリペプチド対する抗体は、グリア細胞の細胞分裂治療用インヒビターとしても利用可能である。
【0024】
本発明は、更に、グリア細胞の細胞分裂を刺激する方法も提供し、該方法は、グリア細胞を以下の式で定義されるポリペプチドに接触させる工程を有する、即ち、
WYBAZCX
ここで、WYBAZCXは、図31に示されるアミノ酸配列から成り(配列認識番号136〜139,141〜147,160,161)、Wは、ポリペプチドセグメントFを有するかあるいは有さず、Yは、ポリペプチドセグメントEを有するかあるいは有さず、Zは、ポリペプチドセグメントGを有するかあるいは有さず、そして、ここにXはポリペプチドセグメントC/D HKL,C/D H,C/D HL,C/D D,C/D’HL,C/D’HKL,C/D’H,C/D’D,C/D C/D’HKL,C/D C/D’H,C/D C/D’HL,C/D C/D’D,C/D D’H,C/D D’HL,C/D D’HKL,C/D’D’H,C/D’D’HL,C/D’D’HKL,C/D C/D’D’H,C/D C/D’D’HL,またはC/D C/D’D’HKLを含む。
【0025】
本発明は、更に、グリア細胞分裂誘発因子を合成する方法に関し、この方法は、上記定義の変成ホスト細胞を、本発明のDNA配列の発現を許容する条件下において培養することから構成される。
【0026】
本発明のペプチドは、薬用又は獣医用としての薬用調合物質又は獣医療調合物質の製造に利用可能である。オプションとして、前記調合物質は、許容可能な希釈液、キャリア又は賦形薬との併用で、及び/又は単位投与薬の形態として使用可能である。
【0027】
グリア細胞を生体内又は生体外においてグリア細胞分裂誘発物質としての上述のポリペプチドと接触させることによってグリア細胞の細胞分裂を刺激する方法も本発明の一態様を構成するものである。有効量の前述のポリペプチドを投与することによって脊椎類(好ましくは哺乳類、更に好ましくはヒト)においてグリア細胞細胞分裂効果を作り出す方法も本発明の一部である。
【0028】
前述のポリペプチドを使用して様々な疾病及び障害を治療する方法も本発明の一部である。例えば、前述のポリペプチドによって、神経疾病又は障害の治療又は予防方法を行うことができる。更に、前記ポリペプチドに対して敏感であるか又は反応する種類の細胞における神経系の病態生理学的症状を予防又は治療する方法も、本発明の一部を構成する。
【0029】
更に、本発明は、末梢神経の損傷、中枢神経系における神経損傷、神経性障害、末梢又は中枢神経系における脱髄、シュワン細胞の損傷又は消失、乏突起膠細胞、小グリア細胞、星状膠細胞に関連する病状の治療方法にも関する。例えば、知覚神経繊維又は運動神経繊維の神経障害や、神経変成性障害の治療が含まれる。これらのいずれの場合においても、その治療方法は、有効量の前記ポリペプチドを投与する工程から成る。
【0030】
本発明は、更に、有効量の前記ポリペプチドを投与することによって、神経の再生及び/又は修復する方法にも関する。そのような薬品は、前記ポリペプチドを薬剤的に有効なキャリアと共に投与することによって得られる。
【0031】
本発明は、薬品の製造における前記ポリペプチドの使用にも関する。
【0032】
本発明は、更に、前記定義のポリペプチドの以下に記載するような使用を含む。
・選択的に治療又は診断目的に利用可能な抗体を生成するために哺乳動物を免疫化するのに利用すること。
・前記ポリペプチドのレセプター結合特性に対応するレセプター結合特性を有する分子を同定したり定量化したりする競合アッセイに利用すること、及び/または、
・前記ポリペプチドに特異的に結合可能なレセプターとともにこのポリペプチドにサンプルを接触させて、該ポリペプチドに対する結合の競合阻害を検出する目的に利用すること、
・例えば、アフィニティークロマトグラフィ等のアフィニティー分離プロセスに使用して対応のレセプターを分離するのに利用すること。
【0033】
本発明は、更に、グリア細胞腫瘍の予防又は治療方法にも関する。この方法は、前記ペプチドによって定義される因子の結合を抑止する物質の有効量を投与することから成る。
【0034】
更に、本発明は、以下に記載するものをグリア細胞に適用することによってグリア細胞細胞分裂誘発活性を刺激する方法にも関する、即ち、
・MDA−MB231ヒト胸細胞ラインから分離された30kDのポリペプチド因子、又は、
・ラットI−EJ形質転換線維芽細胞系からグリア細胞に分離された35kDのポリペプチド因子、又は、
・SKBR−3ヒト胸細胞系から分離された75kDのポリペプチド因子、又は、
・ラットI−EJ形質転換線維芽細胞系から分離された44kDのポリペプチド因子、又は、
・活性化マウス腹膜マクロファージから分離された25kDのポリペプチド因子、又は、
・MDA−MB231ヒト胸細胞から分離された45kDのポリペプチド因子、又は、
・ATL−2ヒトT−細胞ラインからグリア細胞に分離された7ないし14kDのポリペプチド因子、又は
・ウシ腎臓細胞から分離された25kDのポリペプチド因子、又は、
・脳から分離された42kDのポリペプチド因子(ARIA)。
【0035】
本発明は、更に、生体内及び生体外でのグリア細胞の細胞分裂誘発の刺激用の、それぞれ図38ないし43において配列認識番号154〜159によって示されEGFL1、EGFL2、EGFL3、EGFL4、EGFL5、EGFL6ポリペプチドの利用方法にも関する。
【0036】
本発明には、更に、グリア細胞の細胞分裂誘発を刺激する図45のその配列を示すGCF−IIポリペプチドの投与も含まれる。
【0037】
本発明の他の一態様は、上記のペプチドを、シュワン細胞を刺激して科学的用途又は治療用途に採取可能な成長因子を生成させる使用にも関する。
【0038】
更に、ここに記載のペプチドは、再髄鞘形成が必要な、例えばMS等の疾患の治療のための、中枢グリア細胞の増殖及び再髄鞘形成誘発に利用することも可能である。
【0039】
本発明の更に別の態様において、ここに記載の新規なポリペプチドは、アセチルコリンレセプターの合成を刺激するのに利用できる。
【0040】
上述のように、本発明は、公知の因子とは異なった、ウシやヒトを含むほ乳類源から得られる新規なグリア成長因子を提供するものである。これらの因子は、ウシ胎仔血漿(FCP)をバックグラウンドとして、シュワン細胞に対して細胞分裂誘発活性を有する。本発明は、更に、これらの因子の調合方法と、これら及びその他の因子の活性を定義する改良方法も提供するものである。これら因子の治療への適用も本発明の重要な一態様である。
【0041】
従って、本発明の重要な特徴は以下の通りである。
(a)ウシ胎仔血漿の存在下においてグリア細胞分裂誘発活性、特に、シュワン細胞分裂誘発活性を有し、分子量が約30kDないし約36kDの範囲で、そのアミノ酸配列内に、以下に記載のペプチド配列の一つ又は複数を有する基礎(ベーシック)ポリペプチド因子、

(b)ウシ胎仔血漿の存在下において、グリア細胞の細胞分裂誘発、特に、シュワン細胞の分裂誘発を刺激する基礎ポリペプチド因子であって、分子量が約55kDないし約63kDの範囲であり、そのアミノ酸配列内において以下のペプチド配列のうちの一つ又は複数のものを含む、
分子量が小さなポリペプチド因子と、分子量が大きなポリペプチド因子とから得られる上記の新規なペプチド配列も、それ自身本発明の権利の一部を構成するものである。これらの配列は、本発明のポリペプチド因子について、広範囲の様々な種からこのような因子(又は対応の遺伝子配列)を調査し、分離し、あるいは合成し、又は、このような因子を遺伝子組替え技術によって合成するためのプローブ源として有用であり、更には、対応する抗体、好ましくはモノクローナル抗体、であってそれ自身が有用な研究手段であり、また治療物質としての可能性も有する抗体の、従来技術による合成にも有用である。本発明は、更に、本発明の新規なペプチド配列のための前記の方法によって得られる、遺伝子配列又はそのフラグメントをコード化する分離グリア細胞細胞分裂誘発活性にも関する。
【0043】
本発明における高度に精製された因子から得られる短いペプチドの使用により、更に別の配列も特定することが可能になった(下記の例参照)。
【0044】
従って、本発明は、更に、グリア細胞細胞分裂誘発活性を有し、以下に記載するDNA配列によってコード化されるアミノ酸配列を備えたポリペプチド因子も含む。即ち、
(a)図28A、28B又は28Cのいずれかにおいてそれぞれ配列識別番号133〜135で示されるDNA配列、
(b)図22において、配列識別番号89によって示されるDNA配列、
(c)図28Aにおいて配列識別番号133によって示される配列のヌクレオチド281〜557によって表されるDNA配列、
(d)前記(a)、(b)又は(c)によって、いずれかのDNAとハイブリダイゼーション可能なDNA配列。
【0045】
本発明は、更に、上述の配列に対して60%以上、好ましくは80%以上の相同同一性を有する配列を含む。
【0046】
本発明は、特定のハイブリダイゼーション条件に限定されるものではないが、以下の実験記録は、便宜的に従われるガイダンスを提供するものである。
【0047】
DNAプローブは、ショワルター(Schowalter)及びゾマー(Sommer)(Anal.Biochem.,177:90−94,1989)に従い、ニックトランスレーションあるいはPCR反応によって、高比活性(約108ないし109 32Pdmp/μgに標識化して、G−150セファーデックス(Sephadex)カラムでの脱塩によって精製できる。プローブは、変性し(沸騰水中で10分間、その後、冷水に漬ける)、次に、106dpm32P/mlにて10%の硫酸デキストランを含有する80%のバッファB(ポリビニルピロリジン2g、Ficoll−400 2g,ウシ血清アルブミン2g,IM Tris HCL(ph7.5)50ml,NaCl 58g,ピロ燐酸塩ナトリウム1g、ドデシル硫酸ナトリウム10g、H2O 950ml)のハイブリダイゼーション溶液に加え、60℃で一晩(約16時間)培養することができる。次に、前記フィルターを、60℃にて、先ず、バッファB中で15分間、次に、2XSSC、0.1%SDS中で20分間洗浄を3回、IXSSC、0.1%SDS中での20分間の洗浄を1回行うことによって洗浄することができる。
【0048】
その他の態様において、本発明は、以下記載のものを提供する、
(a)ウシ下垂体物質から得られた場合には、還元状態であるか否かに関わらず、下記の分子量標準を使用したSDSポリアクリルアミドゲル電気泳動において、約30kDないし約36kDの分子量が観察される基礎ポリペプチド因子:
リゾチーム(鶏卵白)14,400
大豆トリプシンインヒビター21,500
カルボニックアンヒドラーゼ(ウシ)31,000
卵白アルブミン(鶏卵白)45,000
ウシ血清アルブミン66,200
フォスフォリラーゼB(ラビット筋肉)97,400
そして、当該因子は、ウシ胎仔血漿の存在下において、ラットシュワン細胞の分裂を含むグリア細胞分裂誘発活性を有し、逆相HPLCを使用して分離した場合、4℃で0.1%のトリフルオロ酢酸中での10週間の培養後においてその活性の少なくとも50%を保持する。そして、
(b)ウシ下垂体物質から得られた場合に、非還元状態において、下記の分子量標準を使用したSDSポリアクリルアミドゲル電気泳動において、約55kDないし約63kDの分子量が観察される基礎ポリペプチド因子:
リゾチーム(鶏卵白)14,400
大豆トリプシンインヒビター21,500
カルボニックアンヒドラーゼ(ウシ)31,000
卵白アルブミン(鶏卵白)45,000
ウシ血清アルブミン66,200
フォスフォリラーゼB(ラビット筋肉)97,400
【0049】
そして、当該因子のヒトの等価物質は、ここに記載のDNAクローンGGF2HBS5によってコード化され、また該因子は、ウシ胎仔血漿の存在下において、ラットシュワン細胞の分裂を含むグリア細胞分裂誘発活性を有し、逆相HPLCを使用して分離した場合、4℃で0.1%のトリフルオロ酢酸中での10週間の培養後においてその活性の少なくとも50%を保持する。
【0050】
尚、便宜上、本発明の低い分子量の因子と高い分子量の因子とを、それぞれ、”GGF−I”、”GGF−II”と称する。前記GGF−IIという表示は、GGF−IIタンパク質由来のペプチド配列データで分離されたすべてのクローン(即ち、GGF2HBS5,GGF2BPP3)に使用される。
【0051】
尚、前述の分子量範囲は、正確なものではなく、特定のポリペプチド因子のソースによって僅かに変化するものである。たとえば、別のソースからの物質において約10%の変化は不可能であろう。
【0052】
本発明の別の重要な特徴は、グリア細胞細胞分裂誘発活性を有するポリペプチドをコード化するDNA配列であって、以下のDNA配列である、
(a)図28A,28B又は28Cのいずれかにおいてそれぞれ配列識別番号133〜135で示されるDNA配列、
(b)図22において、配列識別番号89によって示されるDNA配列、
(c)図28Aにおいて配列識別番号133によって示される配列のヌクレオチド281〜557によって表されるDNA配列、
(d)前記(a),(b)又は(c)により、他のDNAとハイブリダイズ可能なDNA配列。
【0053】
本発明の別の態様は、グリア細胞因子とp185erbB2リガンドタンパク質とが前記同じ遺伝子によってコード化されるという事実を利用するものである。この遺伝子から様々なメッセンジャRNAスプライシング変異体が由来し、これらの生成物質の多くが、p185erbB2結合及び活性を示す。前記(GGF−II)遺伝子生成物のいくつかを使用して、シュワン細胞細胞分裂誘発活性が示された。本発明は、前記GGF/p185erbB2リガンド遺伝子のすべて公知の生成物(前記参考文献に記載されている)のシュワン細胞細胞分裂誘発物質としの利用を提供するものである。
【0054】
本発明は、更に、その他のこれまで自然には分離されていないグリア成長因子遺伝子のスプライシング変異体にも関する。図30は、ポリメラーゼ連鎖反応実験(逆転写RNAにおける)とcDNAクローン(ここに記載)の分析、更に、p185erbB2リガンドをコード化する配列として公開されているもの(ペレス(Peles)他,Cell 69:205(1992)及びウェン(Wenn)他,Cell 69:559(1992))からのスプライシングの公知のパターンを示している。これらのパターンは、ここに記載のその他のパターンとともに、存在する可能なスプライシング変異体を表すものである。従って、本発明の別の態様は、この遺伝子から由来の新規なタンパク質因子をコード化するヌクレオチド配列に関する。本発明は、更に、これらの因子の合成方法にも関する。これらの新規な因子の治療への適用も本発明の更に別の態様を構成するものである。
【0055】
従って、本発明の他の重要な特徴は以下の通りである、
(a)シュワン細胞の分裂の刺激を含むグリア細胞細胞分裂誘発活性を有する一連のヒト及びウシポリペプチド因子。これらのポリペプチド配列は、図31,32,33及び34において、それぞれ配列識別番号136〜137として示されている。
(b)シュワン細胞の分裂の刺激を含むグリア細胞細胞分裂誘発活性を有する一連のポリペプチド因子であって、以下の記載概要に基づいて精製され、特徴付けられるもの、即ち、ルプ(Lupu)他,Sclence 249:1552(1990);ルプ(Lupu)他,Proc.Natl.Acad.Sci USA 89:2287(1992);ホルムズ(Holmes)他,Science 256:1205(1992);ペレス(Peles)他,69:205(1992);ヤーデン(Yarden)およびペレス(Peles),Biochemistry 30:3543(1991);ドバシ(Dobashi)他,Proc.Natl.Acad.Sci.88:8582(1991);デイヴィス(Davis)他,Biochem.Biophys.Res.Commun.179:1536(1991);ビューモント(Beaumont)他,特許出願PCT/US91/03443(1990);グリーン(Green)他,特許出願PCT/US91/02331(1990)、アスディン(Usdin)およびフィッシュバック(Fischbach),J.Cell.Biol.103:493−507(1986);フォールズ(Falls)他,Cold Spring Harbor Symp.Quant.Biol.55:397−406(1990);ハリス(Harris)他,Proc.Natl.Acad.Sci.USA 88:7664−7668(1991);及びフォールズ(Falls)他,Cell 72:801−815(1993)。
(c)シュワン細胞の分裂の刺激を含むグリア細胞細胞分裂誘発活性を有するポリペプチド因子(GGFBPP5)。そのアミノ酸配列は、図32において配列認識番号148によって示され、図32において配列認識番号148によって示されるウシDNA配列によってコード化される。
【0056】
上述の新規なヒトペプチド配列は、図31、32、33、34において、それぞれ配列認識番号136〜150として示され、これらは、天然ソース(適当な繊維組織から得られるcDNAライブラリー)からの完全長の相補性DNA(cDNA)として分離可能であり、あるいは、当業者によって、個々のエクソン(例えば、分離エクソンとして得られる)とのDNA構造物として組立可能である。
【0057】
p185erbB2レセプターに特異的に結合する他の化合物、特に、ペプチドも、本発明においては、グリア細胞細胞分裂誘発物質として利用可能である。候補化合物は、p185erbB2結合によりルーチン的にスクリーニングすることが出来、もしも結合する場合には、ここに記載の方法を使用してグリア細胞細胞分裂誘発活性のスクリーニングを行うことができる。
【0058】
本発明は、活性が大幅に減少したものでない、上記ポリペプチド因子のすべての変成物質及び等価物質も含む。例えば、その活性に大きな悪影響を与えることなくアミノ酸内容又は配列を変化させた改変物質が含まれる。例として、天然タンパク質のEP−A109748に於ける突然変異が開示され、ここでは、生物学的反応には必要の無い該天然の配列中のシステインを中性アミノ酸に置換することによって、不要な二硫化物結合の可能性を避けている。従ってここに含まれる効果及び使用方法の記載は、このような改変物質及び同等物質の効果及び使用方法をも含むものであると理解される。
【0059】
本発明の新規な配列は、組替え技術の利点を新たに開くものである。従って、本発明は、以下の態様を含む。
(a)前記構築物による形質転換後の選択されたホスト細胞のベクター(前記配列の発現を許容すべく制御配列に対して位置決めされている)内の作動読み取り枠に於ける前述のDNA配列からなるDNA構築物(好ましくは、前記制御配列は、例えばTrp等の調節可能プロモーターを有している)。尚、プロモーターと調節配列(あるとすれば)の選択は当業者に於ける選択事項である。
(b)前記DNA配列が前記ホスト細胞において発現できるように上記(a)に定義された構築物を組み込むことによって変成されたホスト細胞−ホストの選択は重要でなく、選択される細胞は、原核生物あるいは真核生物のいずれであってもよく、公知の方法によって遺伝子的に変形して前記構築物を組み込むことが可能である。そして
(c)DNA配列の発現を許容する条件下において、変成されたホスト細胞を培養することからなる上記因子の合成方法。前記条件は、DNA組替え技術の当業者によって実施態様に応じて容易に決定することが可能である。本発明は、この手段によって合成されたグリア細胞細胞分裂誘発物質を含む。
【0060】
該技術において記載されているいずれの因子も、本発明の新規なポリペプチド因子が有する組合せの特性を有していない。
【0061】
前述したように、本発明の因子を特徴付けるシュワン細胞分析は、バックグランドとしてウシ胎仔血漿を使用する。その他すべての点においては、この分析は、ブロックス(Brockes)他,in Meth.Enz.,supraと同じであってよいが、但し、10%のFCSを10%のFCPで置き換える。この分析技術の違いは重要である。というのは、ウシ胎仔血漿において血清由来の因子が不在であることによって(血清に対抗するように)、その他のいくつかの因子からの疑似効果を除去することにより、シュワン細胞に於ける活性を厳密に定義することが可能になるからである。
【0062】
本発明は、更に、上述のポリペプチドを調製する方法を含み、該方法は、タンパク質を得るために脊椎類の脳物質を抽出し、この抽出物をハイドロキシアパタイトHPLCによってクロマトグラフィ精製し、次に、これらのフラクションをSDS−ポリアクリルアミドゲル電気泳動にかけることよりなる。約30kDの分子量が観察されたフラクション及び/又は約55kDないし63kDの分子量が観察されたフラクションを採取する。いずれの場合においても、このフラクションを、下記の分子量標準を使用してSDS−ポリアクリルアミドゲル電気泳動にかける。
【0063】
リゾチーム(鶏卵白)14,400
大豆トリプシンインヒビター21,500
カルボニックアンヒドラーゼ(ウシ)31,000
卵アルブミン(鶏卵白)45,000
ウシ血清アルブミン66,200
フォスフォリラーゼB(ラビット筋肉)97,400
【0064】
分子量の小さなフラクションの場合には、前記SDS−ポリアクリルアミドゲルを、非還元条件下、あるいは還元条件下のいずれかでランさせ、分子量の大きなフラクションの場合には、該ゲルを非還元条件下でランさせる。次に、これらのフラクションの、ウシ胎仔血漿のバックグラウンド下でラットシュワン細胞の分裂を刺激する活性についてテストする。
【0065】
好ましくは、上記方法は、例えば、ウシ下垂体物質からのカルボキシルメチルセルロースクロマトグラフィによって得られた関連フラクションを分離することによって始める。又、好ましくは、SDS−ポリアクリルアミドゲル電気泳動の前に、ハイドロキシアパタイトHPLC、陽イオン交換クロマトグラフィ、ゲル濾過、及び/または逆相HPLCを使用する。この方法の各段階において、ブロックス(Brockes)in Meth.Enz.,supraに一般的手段として記載されている分析、但し、10%FCPを10%FCSに代えた分析、により、放射性ヨウ化デオキシウリジンを標準として組み入れシュワン細胞を使用することによって活性を測定することができる。
【0066】
従って、本発明は、更に、分析対象の物質によって刺激された(もしあれば)グリア細胞におけるDNA合成を評価することに対して、バックグランドとしてウシ胎仔血漿を使用するグリア細胞細胞分裂誘発活性の分析方法を含む。
【0067】
本発明の更に別の態様は、オプションとして、許容可能な希釈液、キャリア又は補形薬との併用で、及び/又は単位投与として使用される、薬用又は獣医用として調合された前述のすべての因子からなる薬用又は獣医療用調合物質に関する。本発明の因子の使用においては、適当な調合又は組成を提供するべく従来の薬学的又は獣医学的慣用手段を利用することが出来る。
【0068】
従って、本発明は、例えば、静脈内、皮下内、筋肉内、眼窩内、眼用、心室内、頭蓋内、嚢内、髄腔内、大槽内、腹膜内、鼻腔内、煙霧、乱刺の各投与そして又、経口、口内、直腸、又は膣への投与等の非経口投与法に適用可能である。
【0069】
本発明の調合物は、更に、本発明のDNAを発現するホスト細胞の患者への移植、あるいはこれら調合物を放出する外科移植によって投与することも可能である。
【0070】
非経口調合物の形態は、溶液や懸濁液であってよく、又、経口投与調合物の形態は、錠剤、カプセル等であり、鼻腔内投与調合物の形態は、粉末、鼻腔点滴薬、エアロゾル等とすることができる。
【0071】
公知の調合方法は、例えば、”Remington’s Pharmaceutical Sciences”に記載されている。非経口投与用調合物は、補形薬として、例えば、滅菌した水または塩水、ポリエチレングリコール等のポリアルキレングリコール、植物性オイル、水素化ナフタレン、生物学的適合性、生物分解性ラクチドポリマ、ポリオキシエチレン−ポリオキシプロピレンコポリマー等を使用して本発明の因子の放出を調節することが可能である。他のこれら因子の非経口投与システムとして可能なものとしては、エチレン−ビニルアセテートコポリマー粒子、浸透ポンプ、移植可能注入システムやリポソーム等がある。吸引用の調合物は、補形薬として、例えば、ラクトースを含有することが可能であり、又は、例えば、ポリオキシエチレン−9−ラウリルエーテル、グリココール酸塩やデオキシコール酸塩等を含有する水溶液にしたり、鼻腔点滴液投与用としては油性溶液としたり、あるいは、ゲル状にしたりすることができる。非経口投与用調合物としては、更に、口内投与用にはグリココール酸塩、直腸投与にはメトキシサリチル酸塩、または膣投与用にはクエン酸等がある。
【0072】
本発明の因子は、単独の活性薬剤として使用したり、あるいは他の活性成分、例えば、神経疾患における神経生存を容易にする他の成長因子や、ペプチダーゼ又はプロテアーゼインヒビター等と組み合わせて使用することができる。
【0073】
本発明の調合物における本発明の因子の濃度は、投与される量な投与経路等の多くの要因に応じて変化する。
【0074】
一般に、本発明の因子は、非経口投与用としては水性生理バッファ溶液中に約0.1ないし10%w/v化合物を入れて提供することができる。一般的な投与量は、体重当り、一日につき、約1mg/kgないし約1g/kgの範囲である。好ましい投与量範囲は、体重当り、一日につき、約0.01mg/kgないし100mg/kgである。この好ましい投与量は、治療対象の病理状態の種類と程度、患者の全体的健康状態、調合物の形態及び投与経路によって異なったものになる。
【0075】
前述のように、シュワン細胞(末梢神経系のグリア細胞)は、本発明の因子が存在することによって刺激され分裂する。末梢神経のシュワン細胞は、ニューロンの支持と、個々の神経線維の周りの髄鞘(ミエリン鞘)の形成に関連している。この鞘は、筋肉への電気パルスと知覚レセプターからの電気パルスの適当な伝達にとって重要である。
【0076】
一次的又は二次的にシュワン細胞と神経線維が損傷を受ける数多くの種類の末梢神経障害が存在する。又、知覚神経と運動神経の双方の神経障害も数多くのものがある(アダムズ(Adams)およびヴィクター(Victor),Principles of Neurology)。これらの神経障害のなかで最も重要なものは、恐らく、糖尿病、多発性硬化症、Landry−Guillian−Barr症候群、腫瘍によって発生する神経障害、及び毒物によって生じる神経障害(これらの内のいくつかは腫瘍の治療に利用される)である。
【0077】
しかしながら、本発明は、神経システムの損傷が、例えば、感染又は怪我等いかなる原因によって生じた場合においても使用できる治療方法と予防方法とを提供するものである。従って、脱髄又はシュワン細胞の損失が存在する神経システムの障害又疾患の治療において本発明の因子を使用することに加えて、これらのグリア細胞成長因子は、末梢神経に対する損傷によって生じた神経系の障害の治療においても有効である。末梢神経に対する損傷の後、シュワン細胞の成長又は再形成によって再生プロセスが誘発され、その後、神経線維がその目標に向かって再成長する。シュワン細胞の分裂を加速することによって、損傷後の再生プロセスを促進することができる。
【0078】
中枢神経系(脳及び脊髄)の損傷又は神経退化疾患の治療にも類似の方法を使用することができる。
【0079】
更に、様々なグリア細胞の腫瘍があるが、それらの内で最も一般的なものは、恐らく、神経線維腫であり、これはグリア細胞の過剰成長によって生じる斑状小腫瘍である。又、いくつかのシュワン細胞腫瘍においてGGFに非常に類似した活性物質が見られることも知られており、従って、それらのレセプターに対する本発明のインヒビター活性によってグリア腫瘍を治療することが出来、これは前述ように、因子のレセプターへの結合を阻害するのに有効量の物質を投与することからなる。
【0080】
一般に、本発明は、因子−感受性又は因子−反応性細胞が関連する神経系の病態生理学的症状の予防又は治療に本発明のポリペプチド因子を使用することを含む。
【0081】
本発明のポリペプチド因子は、更に、標準的技術に従って、モノクローナル抗体等の抗体を作る免疫源としても利用可能である。このような抗体も本発明の範囲に含まれる。又、これらの抗体を、治療や診断の目的の為に利用することも可能である。従って、恐らく因子が異常なレベルであることに関連する状態を、このような抗体を利用して追求することができる。標準的方法により単離されたサンプルの分析を利用し、生体外技術が使用できる。又、腫瘍画像化技術を利用して抗体を、例えば、放射性アイソトープで標識化し、体外から画像化できる画像化方法も、使用可能である。
【0082】
本発明は、更に、前記因子を生体内(in vivo)又は生体外(in vitro)でグリア細胞細胞分裂誘発物質として使用することと、そのような用途としての因子とを含む。従って一つの具体的実施例は、有効量の本発明の因子を投与することによって、脊椎類においてグリア細胞細胞分裂誘発効果を生成する方法に関する。そのような方法の好適な実施例は、神経システム疾患又は障害の治療方法又は予防方法である。
【0083】
本発明の更に別の態様は、本発明の因子を、薬品の製造、好ましくは、神経疾患又は障害、あるいは神経再生又は修復のための薬品の製造に使用することである。
【0084】
更に、本発明は、本発明の因子の、前記ポリペプチドに対応のレセプター結合特性を有する分子を同定、あるいは定量する競合アッセイへの利用も含む。これらのポリペプチドは、ラジアイソトープによって標識化してもよい。競合アッセイは、関連するレセプターの拮抗薬と作用薬の両方を同定することができる。
【0085】
別の態様において、本発明は、それぞれの対応レセプターを分離するために、例えばアフィニティークロマトグラフィ等のアフィニティー分離プロセスにおける本発明の因子の使用を提供する。このような特定のタンパク質に対応するレセプターの分離のためのプロセスは、当該技術において公知であり、本発明の因子においても数多くの技術が適用可能である。例えば、IL−6とIFNとに関して、ノヴィック,ディ(Novick,D.)他,J.Chromatogr.(1990)510:331−7が挙げられる。ゴナドトロピン放出ホルモンに関しては文献は以下のものである:ハザム,イー(Hazum,E.),J.Chromatogr.(1990)510:233−8。またG−CSFに関しては、フクナガ,アール(Fukunaga,R.)他,J.Biol.Chem.,265:13386−90がある。IL−2に関しては、スマート,ジェイ,イー(Smart,J.E.)他,(1990)J.Invest.Dermatol.,94:158S−163Sがあり、ヒトIFN−ガンマに関しては、ステファノス,エス(Stefanos,S.)他,(1989)J.Interferon Res.,9:719−30がある。
【0086】
図面の簡単な説明
先ず、図面について説明する。
図面
図1ないし8は、例1に関し、これらについて簡単に説明する。
図1は、カルボキシルメチルセルロースクロマトグラフィからの生成物のプロフィール、
図2は、ハイドロキシアパタイトHPLCからの生成物のプロフィール、
図3は、モノ S FPLCからの生成物のプロフィール、
図4は、ゲル濾過処理FPLCの生成物のプロフィール、
図5及び6は、逆相HPLCからの二つの部分的に精製されたポリペプチド生成物のプロフィールを示す、
図7及び8は、ウシ胎仔血清又はウシ胎仔血漿のバックグランドを使用した逆相HPLCからのGCF−II及びGCF−IIの投与量−反応曲線を示す、
図9ないし12は、GCF−IとGCF−II,配列認識番号1〜20、22〜29、32〜53及び169から由来のペプチド配列を示し(後記の例2参照)、図10及び12は、特に、新規な配列を示す、
図10において、パネルAは、縮重オリゴヌクレオチドプローブと縮重PCRプライマーとを構成するのに使用するGCF−Iペプチドの配列がリストされている(配列認識番号20,1,22〜29及び17)。パネルAのこれらの配列の内のいくつかは、合成ペプチドの構成にも使用された。パネルBは、縮重プローブと縮重PCRプライマーの構成には短すぎた(6アミノ酸以下)新規なペプチドの配列のリストである(配列認識番号17及び52)、
図12において、パネルAは、縮重オリゴヌクレオチドプローブと縮重PCRプライマーとを構成するのに使用するGCF−IIペプチドの配列がリストされている(配列認識番号45〜52)。パネルAのこれらの配列の内のいくつかは、合成ペプチドの構成に使用された。パネルBは、縮重プローブと縮重PCRプライマーの構成には短すぎた(6アミノ酸以下)新規なペプチドの配列のリストである(配列認識番号53)、
図13ないし20は、後記の例3に関し、本発明の因子の細胞分裂誘発活性を示し、
図21ないし28(a,b及びc)は、後記の例4に関し、これらについて以下簡単に説明する、
図21は、図10のパネルA及び図12のパネルAに示された新規なペプチド配列から構成された縮重オリゴヌクレオチド(配列認識番号54−88)のリストである、
図22(配列認識番号89)は、縮重オリゴヌクレオチドプローブ609,650(それぞれ図21,配列認識番号69及び72参照)の結合サイトを含有する組替えウシゲノムファーゼGCF2BG1からの推定ゲノムファーゼGGF2BG1のストレッチを示す。同図は、DNA配列のコード化ストランドと第3読み取り枠の推定されたアミノ酸配列である。因子2(太字)からのペプチド12の配列は、66アミノ酸転写読み取り枠(ヌクレオチド75272)の一部である、
図23は、後部下垂体からのRNAに存在するウシGGF−IIコード下配列のセグメントを分離する実験に使用された縮重PCRプライマー(パネルA,配列認識番号90〜108)とユニークなPCRプライマー(パネルB,配列認識番号109〜119)である、
図24は、図7のパネルA,Bのプライマーのリストを使用したPCR増幅実験において得られた9つの別々の隣接したウシGGF−II cDNA構造体と配列、及び後部下垂体からのRNAを示す。同図の最上ラインは、特徴付けられたcDNA構造体に寄与するコード化配列の略図である、
図25は、GGF2BG1のウシ組替えファーゼの物理的地図である。前記ウシフラグメントは、長さが約20kbであり、ウシGGF−II遺伝子の二つのエクソン(太字)を有している。酵素Xbal,SpeI,Ndel,EcoRI,Kpnl,SstIの制限サイトがこの物理的地図上に示されている。斜線部分は、配列のためにサブクローンされたフラグメントに対応する、
図26は、前記推定ウシGGF−II遺伝子の三つの別の遺伝子生成物の構造の略図である。エクソンはその発見の順番にAないしEとしてリストされている。別のスプライシングパターン1,2及び3は、三つのオーバーラップする推定タンパク質構造体(GGF2BPP1,2,3)を生成し、これらはそれぞれ別の図28A,B,C(下記)に示されている、
図27(配列認識番号120〜132)は、図28A,28B,28C(下記)に示された推定タンパク質配列において同定されたGGF−I及びGGF−IIと、図10及び12にリストされた新規なペプチド配列との比較である。同図は、9つの新規なGGF−IIペプチド配列の内の6つが、これらの推定タンパク質配列に見られることを示している。GGF−I配列に類似の二つのペプチド配列も見られる、
図28A(配列認識番号133)は、図26のスプライシングパターンNo.1から得られるコード化トスランドDNAと推定アミノ酸配列のリストである。前記推定ウシGGF−II遺伝子のこの部分cDNAは、206アミノ酸のタンパク質をコード化する。太字のペプチドは、図10及び12に示したリストから同定されたペプチドである。潜在的グリコシル化サイトがアンダーラインされている(ポリアデニル化信号AATAAAとともに)。
図28B(配列認識番号134)は、図26のスプライシングパターンNo.2から得られるコード化トスランドDNAと推定アミノ酸配列のリストである。前記推定ウシGGF−II遺伝子のこの部分cDNAは、281アミノ酸のタンパク質をコード化する。太字のペプチドは、図10及び12に示したリストから同定されたペプチドである。潜在的グリコシル化サイトがアンダーラインされている(ポリアデニル化信号AATAAAとともに)。
図28C(配列認識番号135)は、図26のスプライシングパターンNo.3から得られるコード化トスランドDNAと推定アミノ酸配列のリストである。前記推定ウシGGF−II遺伝子のこの部分cDNAは、257アミノ酸のタンパク質をコード化する。太字のペプチドは、図10及び12に示したリストから同定されたペプチドである。潜在的グリコシル化サイトがアンダーラインされている(ポリアデニル化信号AATAAAとともに)。
図29は、下記の例6に関し、サザンブロット上の様々なほ乳類DNAに対する推定ウシGGF−II遺伝子配列のクロスハイブリダイゼーション分析のオートラジオグラムである。前記フィルターは、同図にリストされた種からのEcoRI−消化DNA(各レーンにつき5μg)を含有する。前記プローブは、図25の物理的地図によって予測されるウシDNAの4キロベースのフラグメントを含んで、各DNAサンプルにおける単一の強力なバンドを検出する。より力の小さいバンドも観察され、これらは関連DNA配列を示す。他のほ乳類DNAサンプルからの強いハイブリダイゼーションバンドは、これらの種のGGF−II相同体を示すものと考えられる。
図30は、さまざまなスプライシング変異体を示す図である。コード化セグメントは、F,E,B,A,G,C,C/D,C/D’,D,D’,H,K及びLによって示されている。精製タンパク質からのペプチド配列の位置は、”0”によって示されている。
図31(配列認識番号136〜147,160,161)はGGFのコード化セグメントのDNA配列と予想ペプチド配列のリストである。ライン1は、ウシGGFの予想アミノ酸配列のリストであり、ライン2は、ウシGGFのヌクレオチド配列のリストであり、ライン3は、ヒトGGF(ヘレグリン)のヌクレオチド配列のリストであり(ヌクレオチドベースのマッチングは縦線にて示してある)、ライン4は、前記予想ウシ配列とは異なる場合におけるヒトGGF/heregulinの予想アミノ酸配列のリストである。コード化セグメントE、A’及びKは、ウシ配列のみを表す。コード化セグメントD’は、ヒト(ヘレグリン)配列のみを表す。
図32(配列認識番号148)は、BPP5の予想GGF2アミノ酸配列とヌクレオチド配列である。上方ラインは、ヌクレオチド配列であり、下方ラインは、予想アミノ酸配列である。
図33(配列認識番号149)は、GGF2BPP2の予想アミノ酸配列とヌクレオチド配列である。上方ラインは、ヌクレオチド配列であり、下方ラインは、予想アミノ酸配列である。
図34(配列認識番号150)は、GGF2BPP4の予想アミノ酸配列とヌクレオチド配列である。上方ラインは、ヌクレオチド配列であり、下方ラインは、予想アミノ酸配列である。
図35(配列認識番号151〜152)は、二つのGGFペプチド配列(GGF2pp4とGGF2pp5)のヒトEGF(hEGF)との整合を示す。星印は、保存されたシステインを示す。
図36は、GGFの増加に対する、約200kD(抗燐酸化ポリクローナル抗体とともに現像したウエスタンブロット上における200kDバンドの強度)のタンパク質のGGF活性のレベル(シュワン細胞細胞分裂誘発分析)と、チロシン燐酸化を示す。
図37は、図31の配列からのスプライシング変異体のリスト、
図38は、EGFL1(配列認識番号154)の、予想アミノ酸配列を下部に、核酸配列を上部に示す、
図39は、EGFL2(配列認識番号155)の、予想アミノ酸配列を下部に、核酸配列を上部に示す、
図40は、EGFL3(配列認識番号156)の、予想アミノ酸配列を下部に、核酸配列を上部に示す、
図41は、EGFL4(配列認識番号157)の、予想アミノ酸配列を下部に、核酸配列を上部に示す、
図42は、EGFL5(配列認識番号158)の、予想アミノ酸配列を下部に、核酸配列を上部に示す、
図43は、EGFL6(配列認識番号159)の、予想アミノ酸配列を下部に、核酸配列を上部に示す、
図44は、クローンのスケールコード化セグメントマップである。T3は、前記クローンからmRNAを生成するのに使用するバクテリオファージプロモーターを示す。R=隣接のEcoRI制限酵素サイト。5’UTは、5’非翻訳領域を示す。E,B,A,C,C/D’及びDは、コード化セグメントを示す。0=翻訳開始サイト。Λ=ウシEセグメントに対して相同の領域の5’限界(例6参照)、そして、3’UTは3’非翻訳領域を示す。
図45は、GGF2HBS5の予想アミノ酸配列(中間部:配列認識番号170)と、核酸配列(上部:配列認識番号21)とを示す。下部(中間)配列は、GGF−II調製物(図11及び12参照)からのペプチド配列を示す。
図46は、組替えヒト及びウシグリア成長因子のシュワン細胞細胞分裂誘発活性を示すグラフ。
図47は、CHO細胞のならし培地の大きさの異なる部分標本(aliquots)の投与から生じたシュワン細胞増殖活性物質の投与量−反応曲線である。
図48は、GGF2HBS5cDNAクローンを含有するバキュロウィルスによって感染したSF9昆虫細胞によって細胞外培地中に分泌されたシュワン細胞細胞分裂誘発活性物質の投与量−反応曲線、
図49は、GGFペプチド抗体を使用した組替えCHO細胞のならし培地のウェスタンブロット、
図50Aは、陽イオン交換カラムから溶出した組替え(COS細胞が生成)ヒトGGF−II(rhGGF−II)ピークのシュワン細胞増殖活性のグラフであり、
図50Bは、rhGGFIIの特定のペプチドに対して生成されたポリクローナル抗体を使用した組替えGGFIIピークに対する免疫ブロットである、
図51Aは、フラクションごとの陽イオン交換カラムでのrhGGF−II(CHO細胞が生成)の精製を示すグラフ、
図51Bは、図51Aに示されたフラクションとrhGGF−II特異抗体を使用したウエスタンブロットの写真である、
図52は、組替えグリア成長因子で処理したシュワン細胞におけるチロシン燐酸化を示すゲルの写真、
図53は、GGFHBS5,GGFHFB1及びGGFBPP5ポリペプチド(配列認識番号170,171,172)の配列、
図54は、前記CHO細胞−発現ベクターpcDHFRpolyAのマップである。
【0087】
詳細な説明
本発明は、新規なグリア成長因子の分離と精製、及び、これら因子をコード化するDNA配列のクローニングとに関する。本発明の他の組成物は、一連のグリア成長因子を潜在的にコード化できるいくつかの遺伝子スプライシング変異体、特に、GGF2HBS5、特にウシGGF−IIのヒト等価物をコード化する変異体である。GGF’s及びp185erbB2結合タンパク質をコード化する遺伝子が、長さが異なりいくつかの共通のペプチド配列といくつかのユニークなペプチド配列とを有する一連のタンパク質を生み出すRNA転写体であって、様々な大きさとスプライス状態の異なったRNA転写体を生成することは明らかである。これは、ウシ後方下垂体RNA(ここに開示される)、ヒトの乳ガン(MDA−MB−231)(ホルムズ(Holmes)他,Science 256:1205(1992))や鶏の脳のRNA(フォールズ(Falls)他,Cell 72:1−20(1993))から回収可能なスプライシングの異なる諸配列によって証拠付けられている。又、シュワン細胞に対する細胞分裂誘発物質(ここに開示)として、また、前記p185erbB2レセプター(下記参照)のためのリガンドの両方として作用する広範囲のタンパク質によっても証拠付けられている。
【0088】
前記GGFとp185erbB2をコード化する遺伝子が相同であるという事実の更なる証拠は、ヌクレオチド配列の比較から得られる。Science,256(1992),(1205〜1210)ホルムズ(Holmes)他,は、前記レセプタータンパク質p185erbB2に特異的に反応し、いくつかのヒトの悪性腫瘍に関連する45−キロダルトンのヒトタンパク質(ヘレグリン−α)の精製について記載している。ヘレグリン−αをコード化するいくつかの相補性DNAクローンが分離された。ペレス(Peles)他(Cell 69:205(1992))及びウェン(Wen)他(Cell 69:559(1992))は、「neu分化因子」(NDF)と称するタンパク質をコード化するラット細胞から分離された相補性DNAについて記載している。前記NDFcDNAのトランスレーション生成物は、p185erbB2結合活性を有する。アスディン(Usdin)及びフィッシュバック,ジェイ(Fischbach,J.)Cell.Biol.103:493−507(1986);フォールズ(Falls)他,Cold Spring Harbor Symp.Quant.Biol.55:397−406(1990);ハリス(Harris)他,Proc.Natl.Acad.Sci.USA 88:7664−7668(1991);及びフォールズ(Falls)他,Cell 72:801−815(1993)は、レセプタータンパク質p185erbB2と作用する42Kdのグリコプロテインの精製されたこと、また、いくつかの相補性cDNAが分離されたこと(フォールズ(Falls)他Cell 72:801:815(1993))を示している。その他のグループとしては、ルプ(Lupu)他(1992)Proc.Natl.Acad.Sci.USA 89:2287;ヤーデン(Yarden)とペレス(Peles)(1991)Biochemistry 30:3543;ルプ(Lupu)他(1990)Science 249:1552;ドバシ(Dobashi)他(1991)Biochem.Biophys.Res.Comm.179:1536;そしてフアング(Huang)他(1992)J.Biol.Chem.257:11508−11512等が含まれる。
【0089】
他の実施例
本発明は、図31のコード化セグメント(配列認識番号136〜147,160及び161)にほぼ相同のすべてのタンパク質と、自然発生GGFポリペプチドとを含む。更に、対立遺伝子変異体、自然突然変異体、誘発突然変異体、自然発生の核酸に対する高ストリンジェント条件又は低ストリンジェント条件下においてハイブリダイズしたDNAによってコード化されたタンパク質(高ストリンジェント条件及び低ストリンジェント条件の定義に関しては、ここに参照文献として添付するCurrent Protocols in Molecular Biology,John Wiley & Sons,New York,1989,6.3.1−6.3.6を参照)、そして、抗血清によってGGFポリペプチドに特異的に結合したポリペプチド又はタンパク質も含まれる。又、この用語は、図31からの配列を有するGGFポリペプチドを含むキメラポリペプチドも含む。
【0090】
以下の例は、本発明を限定するものではなく、本発明を有用に例示する目的で提供されるものであり、有効な調製技術に関するガイダンスを提供するものである。
【0091】
下記の例3から理解されるように、本発明の因子は、ある範囲のタイプの細胞に細胞分裂誘発活性を示す。線維芽細胞に関する活性は、傷修復能力があることを示し、本発明は、この利用方法も含むものである。上述の調合物及び/又は薬品、及びその製造方法に関連の本発明の一般的記載は、適当な製造物とその使用方法を含むものであると理解されるべきである。これは、線維芽成長因子(FGFs)に対する類似の活性に関する報告に鑑みて、本発明に対し合理的に期待されるものである。例えば、Sporn et al.,「ペプチド成長因子とそのレセプターI」396頁(Baird and Bohlen)の「傷の治癒と繊維組織の修復におけるFGFs」というタイトルを付けられた章を参照。
【0092】
例1
ウシ下垂体からのGGF−IとGGF−IIとの精製
I.因子−CMフラクションの調製
4,000の冷凍全ウシ下垂体(約12kg)を一晩解凍し、先ず水で簡単に洗浄し、次に、Waring Blenderによって各々同量の0.15M硫酸アンモニウムでバッチ処理によりホモジナイズした。このホモジナイズ化物を1.0MのHClでpH4.5に調節し、4,900gで80分間遠心分離機にかけた。上澄液中のすべての脂肪分を、グラスウールを通過させることによって除去した。前記上澄液のpHを1.0MのNaOHを使用して6.5に調整した後、固体の硫酸アンモニウムを添加して36%の飽和溶液となるようにした。数時間の攪拌後、その縣濁液を4,900gで80分間遠心分離機にかけ、沈澱物を取り除いた。グラスウールでのフィルター処理後、更に前記上澄液に固体硫酸アンモニウムを追加し、75%の飽和溶液となるようにし、数時間の攪拌後、これを再び4,900gで80分間遠心分離機にかけた。そのペレットを、約2リットルのpH6.0の0.1M燐酸ナトリウム中にて再縣濁し、3ラ40Lの同じバッファで透析した。透析物の導電性が20.0μジーメンス以下であることを確認した後、毎分2mlの流量で、カルボキシルセルロース(CM−52,Whatman)でパックしたバイオプロセスカラム(120ラ113mm,Pharacia)に負荷した。このカラムを、先ず、2倍量のpH6.0の0.1M燐酸ナトリウムで、次に、2倍量の50mMNaClで、そして最後に2倍量の0.2M NaClで、同じバッファ中にて洗浄した。最終工程中において、10mL(5分間)のフラクションが収集された。フラクション73〜118までをプールし、10倍量のpH6..0の10mM燐酸ナトリウムで2度、透析し、100,000gで60分間の遠心分離によって浄化した。
【0093】
II.ハイドロキシアパタイトHPLC
ハイドロキシアパタイトHPLCは、グリア成長因子の分離において従来使用された技術ではないが、本発明において特に有効であることが判った。
【0094】
前記CM−セルロースクロマトグラフィによって得られた物質を、0.02μmのフィルター(Nalgene)によって濾過し、室温にて、ガードカラム(15×25mm,Biorad)を備えた高性能ハイドロキシアパタイトカラム(50×50mm,Biorad)に負荷し、pH6.0の10mMの燐酸カリウムで平衡させた。室温での抽出を、下記のプログラムされたリニアグラジエントを使用して毎分2mlの流量で行った。
【0095】
6.0mL(3分間)フラクションを、前記グラジエント溶出中に採取した。フラクション39〜45をプールし、10分量の50mM燐酸ナトリウムpH6.0で透析した。
【0097】
III.モノ S FPLC
モノ S FPLCにより、後のゲル濾過のためにより高濃度の物質を作ることが可能となった。
【0098】
前記ハイドロキシアパタイトカラムからのプールされた物質のすべての粒状成分を、調製HR10/10モノ S 陽イオン交換カラム(100×10mm,Pharmacia)にかける前に、10,000gでの浄化スピンによって、60分間で除去し、次に、これを室温下で、毎分1.0mlの流量でpH6.0の50mM燐酸ナトリウムに再平衡させた。これらの条件下において、結合したタンパク質を、下記のプログラムされた一次グラジエントを使用して溶出した。
【0099】
1.0mL(1分間)フラクションを、このグラジエントプログラム中に採取した。フラクション99〜115までをプールした。
【0101】
IV.ゲル濾過FPLC
この工程は、最終精製処理の前に、本発明の二つの因子の分離より始め、濃縮フラクションを作った。
【0102】
この実験の目的のため、調製スペローズ(Superose)12 FPLC カラム(510×20mm,Pharmacia)を、その製造会社の指示に従って充填した。このカラムを標準化するために、製造会社の指示に従って、理論段数測定を行い、9,700理論段数という値を得た。
【0103】
モノ S 溶出物質のプールを、室温において、2.5Mlの部分標本ごとに、予めC18逆相カラム(Sep−pak Millipore)に通過させた50mM燐酸ナトリウム、0.75NaCl pH6.0中で毎分1.0mLの流量でこのカラムに適用した。各サンプルが該カラムに適用された後、35分後から1mL(0.5分間)のフラクションが採取された。各実行(ラン)からフラクション27ないし41(GGF−II)及び42〜57(GGF−I)
までがプールされた。
【0104】
V.逆相HPLC
上記スペローズの12回のランから得られたGGF−1とGGF−IIとを、それぞれ、三つの等しい部分標本に分割した。各部分を、ガードカートリッジ(RP−8,15×3.2mm,Applied Biosystems)によって保護され、40℃に平衡させた、C8逆相カラム(Aquapore RP−300 7μ C8 220×4.6mm,Applied Biosystems)にて、毎分0.5mLの流量で通過させた。これらの条件下において、下記のプログラムによるリニアグラジエントを使用してタンパク質を溶出した。
【0105】
200μL(0.4分間)のフラクションを前記プログラムグラジエントの開始後からの15.2分間でシリコン化チューブ(Multilubeチューブ、Bioquote)内に採取した。
【0107】
VI.SDS−ポリアクリルアミドゲル電気泳動
この工程においては、Bio−Rad Labo−ratories Limited,Watford,Englandのタンパク質分子量標準、低レンジ、カタログNo.161−0304を使用した。実際に使用されたタンパク質とその分子量標準は、前述の通りであった。
【0108】
前記逆相プログラムのランから得たフラクション47〜53(GGF−I)とフラクション61〜67(GGF−II)を、個別にプールした。このプールした物質の7μlを、0.0125M Tris−Cl,4%SDS,20%グリセロール、及びGGF−Iに対しては10%のβ−メルカプトエタノールの等量混合液中で、5分間沸騰させ、4%の濃縮用ゲルを有する11%ポリアクリルアミドLaemmliゲル中に充填し、50Vの定電圧を16時間かけた。このゲルを、次に、固定し、銀染色キット(Amersham)を使用して染色した。これらの条件下において、前記因子は、分子量マーカーを基準として、相対分子量が30,000ないし36,000ダルトン(GGF−I)と55,000ないし63,000ダルトン(GGF−II)の、幾分拡散したバンドとして観察される。前記ゲル染色から、前記逆相プログラム実行からプールされた物質において前記GGF−I(5GGF−IIと同等のレベルで、他のタンパク質が存在することが明らかである。
【0109】
VII.トリフルオロ酢酸中における安定性
トリフルオロ酢酸存在下における、本発明の因子の安定性に関して得られたデータは以下の通りである。
【0110】
GGF−I:0.1%TFAとアセトニトリルの存在下において、前記逆相HPLCから得た物質を、カラムランの完了後12時間以内と、その40℃での10週間の培養後とにおいて分析した。培養後、GGF−Iは、カラム流出後、直接にとって分析した物質の活性の少なくとも50%を保持していた。
GGF−II:0.1%TFAとアセトニトリルの存在下において、前記逆相HPLCから得て、−20℃て保存しておいて物質を、解凍後と、40℃で4日間の培養後とにおいて分析した。培養後、GGF−IIは、解凍直後の物質の活性の少なくとも50%を保持していた。
【0111】
尚、上記研究において使用されたトリフルオロ酢酸濃度は、逆相クロマトグラフィにおいて最も一般的に使用されるものである。
【0112】
VIII.活性分析条件
特に明記の無い限り、すべての作業は37℃で行った。そして図1ないし6に示されるように、各工程での活性は、下記の変更以外には、ブロックス(Brockes)(Meth.Enz.,supra)の技術を使用して測定された。従って、シュワン細胞の調製において、5μMのフォルスコリン(forskolin)を、DMEM(ダルベッコ変法イーグル培地)、FCS及びGGF以外に添加した。分析に使用した細胞は、継代数が10以内の無線維芽細胞シュワン細胞であり、これらの細胞は、トリプシンとともにフラスコから除去され、1ミクロ・ウェル当り3.3千細胞の割合で平底96−ウェル・プレートにプレート化した。
【0113】
[125I]IUdRを、テスト溶液の添加後の最後の24時間後に添加した。バックグラウンド(非刺激)の各分析に対する関与は、100cpm以下であり、最大の組込みは、シュワン細胞のバッチと継代数とに応じて、20ないし200倍(fold)であった。
【0114】
上述の逆相HPLCから得られるGGF−I及びGGF−IIフラクションの場合、各因子に対する一本の曲線について、全く同じ前述の方法を使用して、二本の投与量−反応曲線が得られた。そして、この分析実験において上記方法は、各因子についての他の曲線を得るために、ウシ胎仔血漿をウシ胎仔血清に代えた点のみ変更された。その結果は、図7及び8に示されている。
【0115】
例2
精製GGF−IとGGF−IIのアミノ酸配列
高度に精製したウシ下垂体GGF−I及びGGF−IIを使用してアミノ酸配列分析研究を行った。これらの配列の記載には従来式の単一文字コードを使用した。ペプチドは、還元されそしてカルボキシメチル化されたサンプルについて、リシルエンドペプチダーゼとプロテアーゼV8による消化物として得られた。特に前記GGF−IIのリシルエンドペプチダーゼの消化は、11%SDS−PAGE(前記マーカーに対するMW)の55〜−65RD領域において行った。
【0116】
GGF−Iに関して全部で21のペプチド配列(図9、配列認識番号1〜20,169参照)が得られ、その内の12のペプチド(図10、配列認識番号1,22〜29,17,19及び32)は現在のタンパク質データベースには存在しないものであり、従って、新規な配列である。GGF−IIに関して全部で12のペプチド配列(図11)配列認識番号33〜44参照)が得られ、その内の10のペプチド(図12、配列認識番号45〜53)は現在のタンパク質データベースには存在しないものであり、従って、新規な配列である(例外は、ペプチドGGF−II 06であり、これは多くのタンパク質においても同じ配列を示しているが、これらはその残基の数が小さいことから恐らく重要ではない)。これらの新規な配列が、GGF−IとIIの真のアミノ酸配列の部分に対応するものである可能性は極めて高い。
【0117】
特に注目されるのはGGF−I 07とGGF−II 12とであって、これらは明らかに互いに密接に関連している。その類似性は、これらのペプチドの配列がほぼ確実に指定されたGGF類のそれであることと、汚染タンパク質由来である可能性が非常に低いこととを示すものである。
【0118】
更に、ペプチドGGF−II 02において、配列X S Sは、Xによって示す位置におけるアスパラギンのN−結合の炭水化物部分(moiety)の存在と一致している。
【0119】
一般に、図9及び11において、Xは、サイクル中に同じ大きさの信号が一つ以上存在したか、あるいは、全く信号が無かったことによって一つの位置も確実に呼び出すことができなかったシーケンスサイクルを表す未知の残基を示している。星印は、最後に呼び出したアミノ酸が、そのペプチドに存在する最後のアミノ酸に対応するペプチドを示す。その他のタンパク質において、最後に呼び出したアミノ酸の後の信号強度では、そのペプチドの末端まで呼び出しを継続するのには不十分であった。右側のカラムは、NBRFとEMBL配列データベースを分析するためにGCGパッケージFASTA及びTFASTAプログラムを使用して行ったコンピュータデータベースサーチの結果を示している。このカラムのタンパク質の名称は、その配列の一部の一致を示し、呼び出されたペプチドアミノ酸配列は最大2つのミスマッチを許容するものであった。疑問符(?)は、許容された3つのミスマッチを示している。使用された略記は以下の通りである。
【0120】
HMG−1 高移動性グループタンパク質−1
HMG−2 高移動性グループタンパク質−2
LH−アルファ 黄体形成ホルモンアルファサブユニット
LH−ベータ 黄体形成ホルモンベータサブユニット
【0121】
例3
精製GCF−IとGCF−IIの細胞分裂誘発活性
GCF−IとGCF−IIとの両方を含有する高度に精製したサンプルの細胞分裂誘発活性の研究を、定量方法により行った。この方法は、単一のミクロ培養による、DNA合成、細胞組織形態、細胞数、細胞アンチゲンの発現の評価を可能にするものである。この技術は、以前にミュール(Muir)他,Analytical Biochemistry 185,377−382,1990によって報告された方法を変更したものである。主な変更点は、1)非被覆ミクロタイター(microtiter)プレートの使用、2)一ウェル当りの細胞数、3)10%ウシ胎仔血清(FCS)に代えての5%の胎児ウシ血漿(FBP)の使用、そして4)培養に同時に添加された細胞分裂誘発物質と臭化デオキシウリジン(BrdU)の存在下における培養時間、である。更に、細胞の損失を避けるために、前記細胞の単層は、固定前において洗浄しなかった。そして、モノクローナル・マウス抗−BrdU抗体とペルオキシダーゼ接合ヤギ抗−マウス免疫グロブリン(IgG)抗体とを、分析の感度を高めるために、2倍にした。ラットの座骨神経シュワン細胞ように最適化した分析も、前記細胞培養条件に対して適当な変更を施した後に、いくつかの細胞系に使用した。
【0122】
I.細胞分裂誘発テスト
第1日目に、精製したシュワン細胞を、5%EBP/ダルベッコ変法イーグル培地(DMEM)(5,000細胞/ウェル)中で、非被覆の96ウェルのプレートにプレート化した。第2日目に、GGF又はその他のテスト因子を、10μmの最終濃度のBrdUとともに、前記培養物に添加した。48時間後(第4日目)、BrdU組み込みを、前記培地を吸引することによって終わらせ、室温で20分間、70%エタノールを200μl/ウェル添加し、細胞を固定した。次に、これらの細胞を、水及び37℃で10分間、100μlの2N HClでの培養によって変質させたDNAで洗浄した。吸引後、残りの酸を、前記ウェルを0.1Mホウ酸塩バッファpH9.0で満たすことによって中和し、細胞を燐酸緩衝食塩水(PBS)で洗浄した。次に、細胞を、50μlのブロッキングバッファ(0.1%のトリトンX100と2%の正常ヤギ血清とを含有するPBS)で37℃にて15分間処理した。吸引後、モノクローナル・マウス抗−BrdU抗体(ダコ・コーポレイション(Dako Corp.),カリフォルニア州,サンタ・バーバラ)(50μl/ウェル,ブロッキングバッファ中で1.4μg/mlに希釈)を添加し、37℃で2時間培養した。非結合抗体を、0.1%トリトンX−100を含有のPBS中において3回洗浄して除去し、ペルオキシダーゼ−結合ヤギ・抗マウスIgG抗体(ダコ・コーポレイション(Dako Corp.),カリフォルニア州,サンタ・バーバラ)(50μl/ウェル,ブロッキングバッファ中で2μg/mlに希釈)を添加し、37℃で1時間培養した。PBS/トリトンでの3回の洗浄と、PBS中での最終すすぎ後、ウェルは、0.05%の溶解性クロモゲンO−フェニレンジアミン(OPD)と、0.02%のH2O2とを含有する100μl/ウェルの50mMのpH5.0燐酸/クエン酸塩バッファを受けた。室温で5〜20分後、各ウェルから80μlを、2N硫酸を40μl/ウェル含有するクリーンなプレートにピペットで移すことによって、前記反応をに終結させた。490nmにおける吸光はプレートリーダー(Dynatech Labs)を使用して記録した。前記細胞単層を含有する分析プレートを、PBSで2回洗浄し、100μl/ウェルの基質ジアミノベンジジン(DAB)を加えてBrDU−DNAについて免疫細胞化学的に染色し、0.02%のH2O2とを添加することによって不溶性生成物とした。10〜20分後、前記染色反応を、水で洗浄することによって終結させ、倒立顕微鏡(inversed microscope)を使用してBrdU−ポジティブ細胞核を観察計数した。場合により、ネガティブ核を0.001%のトルイジンブルーで対比染色し、上述と同じように計数した。
【0123】
II.細胞分裂誘発分析に使用した細胞系
Swiss 3T3線維芽細胞:Flow Labsからの細胞を、10%CO2の空気中で、加湿雰囲気下、37℃で、10%のFCS、ペニシリン、ストレプトマイシンを補足したDMEM中に保持した。細胞を、2日毎に供給又は二次培養した。細胞分裂誘発分析のために、細胞を、完全培地にて5,000細胞/ウェルの濃度でプレート化し、細胞が融合性を得て、静止状態になるまで1週間培養した。血清を含有する培地を除去し、細胞単層を、無血清培地で2回洗浄した。細胞分裂誘発物質を含有する100μlの無血清培地と、10μMのBrdUとを各ウェルに添加し、48時間培養した。GGFと血清又はPDGF(ホジティブコントロールとして)に対する投与量反応を行った。
【0124】
BHK(ベビーハムスター腎臓)21 C13線維芽細胞:動物細胞培養ヨーロッパ保存機関(ECACC)からの細胞を、5%CO2空気中、加湿雰囲気下、37℃で、5%のFCS、ペニシリン、ストレプトマイシンを補足したグラスゴー変法イーグル培地(GMEM)中で保持した。細胞を、2ないし3日毎に供給又は二次培養した。細胞分裂誘発分析のために、細胞を、完全培地にて、2,000細胞/ウェルの濃度で、24時間プレート化した。血清を含有する培地を除去し、無血清培地で2回洗浄した後、100μlの、0.1%のFCSを含有するGMEMあるいはGMEMのみで、置換した。ホジティブコントロールとしてGGFとFCS又はbFGFとを、10μMのBrdUとともに、添加し、48時間培養した。次に、細胞培養のシュワン細胞を、前述の方法で処理した。
【0125】
C6ラットグリオマ細胞系:継代39から得た細胞を、10%CO2空気中、加湿雰囲気下、37℃で、5%のFCS、ウマ血清(HS)、ペニシリン、ストレプトマイシンを含有したDMEM中で保持した。細胞を、3日毎に供給又は二次培養した。細胞分裂誘発分析のために、細胞を、完全培地にて2,000細胞/ウェルの濃度でプレート化し、24時間培養した。次に、培地を、無血清培地中にて洗浄した後、DMEMと0.1%のFCSを含有するF12の1:1の混合物にて置換した。次に、GGF,FCS及びαFGFに対する投与量反応を行い、細胞を、他のタイプの細胞に関して前述した方法でELISAを通じて処理した。
【0126】
PC12(Rat Adrenal Pheo−chromocytoma細胞):ECACCからの細胞を、5%CO2空気中、加湿雰囲気下で、37℃でコラーゲン被覆フラスコ内にて、10%のFCS、ペニシリン、ストレプトマイシンを補足したRPMI1640中で維持した。細胞を、3日毎に培地の80%を置換することによって、供給した。細胞分裂誘発分析のために、細胞を、完全培地にて3,000細胞/ウェルの濃度でコラーゲン被覆プレート(50μl/ウェルコラーゲン、ヴィトロゲン・コラーゲン・コーポレイション(Vitrogen Collagen Corp.)、1:50に希釈、37℃で30分間)にプレート化し、24時間培養した。次に、前記培地を、新たなRPMIのみか、あるいは、1mMのインシュリンを含有したもの、あるいは、1%のFCSによって置換した。ポジィティブコントロールとしてのFCS/HS(1:2)及びGGFとに対する投与量反応を前述のように行った。48時間後、細胞を固定して、ELISAを前述したように行った。
【0127】
III.細胞分裂誘発分析の結果:
この例におけるすべての実験は、セファロース12クロマトグラフィー精製工程(例1のセクションD参照)からの、GGF−1とGGF−II(GGFs)との混合物を含有する高度に精製したサンプルを使用して行った。
【0128】
先ず、BrdU組み込み分析によって得られた結果を、J.P.Brockes(Methods Enzymol.147:217,1987)に記載の、分裂細胞のDNAへの[125]I−UdR組み込みに基づくシュワン細胞の古典的な細胞分裂誘発分析と比較した。
【0129】
図13は、同じ細胞培養条件(5,000細胞/ウェル5%FBP/DMEM中、GGFs又の存在下で48時間培養)で行われた二つの分析によって得られた結果を比較して示したものである。同図に明らかに示されているように、これらの結果は比較可能であるが、BrdU組み込み分析の方が、グラフにおいて曲線が左側、即ち、GGFSの低い濃度の方へシフトしていることに示されるように、僅かに感度が高い。
【0130】
「細胞分裂誘発テスト方法」と題された章に記載されているように、免疫反応性BrdU−DNAを、前記OPDペルオキシダーゼ反応の溶解生成物強度を読み取ることによって定量化した後、細胞単層を含有する元の分析プレートを、第2の反応をさせて、非溶解性DAB生成物を得ることができ、これはBrdUポジティブ細胞核を染色する。前記マイクロ培養を、次に倒立顕微鏡で調べ、細胞の形態と、BrdU−ポジティブ及びネガティブ細胞核の数を観察することができる。
【0131】
図14a及び図14bにおいて、490nmでの吸収の読み取りによって評価されたBrdU−DNAの免疫活性が、同じ培養内における、BrdU−ポジティブ細胞核の数と、ウェル当りのBrdU−ポジティブ細胞核の百分率と比較されている。標準偏差は、10%以内であった。二つの評価方法は、GGFsの最大投与量における値の間の非常に良好な相関関係と不一致とが、BrdU−ポジティブとして検出された細胞におけるDNA合成の程度の違いによって説明可能であることを示している。
【0132】
従って、BrdU組み込み分析は、前記(125)I−UdR組み込み分析との比較した場合、シュワン細胞に対するポリペプチドの生物学的活性についての別の有用な情報を提供するものである。例えば、図15に報告されるデータは、GGFsがシュワン細胞に作用して、DNA合成を誘発することができるが、より少ない投与量においては、48時間後においてマイクロ培養中に存在するネガティブ細胞の数を増やすことを示している。
【0133】
次に、前記分析を、起源の異なるいくつかの細胞系に使用した。図16において、GGFsに対する、シュワン細胞とSwiss線維芽細胞との細胞分裂誘発反応が比較されており、3T3線維芽細胞においては弱い反応しか得られなかったにもかかわらず、これらの培養中においていくつかのはっきりとしたBrdU−ポジティブ細胞核が検出された。コントロール培養を、FCS又はヒト組替えPDGFの複数の投与量の存在下において平行に実行したところ、これら細胞が適当な刺激に対して反応可能であることが判った(図示せず)。
【0134】
線維芽細胞のGGFsに対する反応性を、更に、BHK 21 C13細胞系を使用して調べた。これらの腎臓から得た線維芽細胞は、接触阻止反応を示さず、あるいは、融合性状態の場合に静止状態には到らない。従って、細胞の生存力を損なわない範囲で、バックグラウンドの増殖が非常に低くなるように実験条件を調節した。GGFsは、図17及び18に示すように、BHK 21 C13細胞において非常に顕著な細胞分裂誘発活性を有する。図17は、0.1%のFCSの存在下においてGGFSによって刺激されたBHK21 C13細胞による、BrdUのDNAへの組み込みを示している。FCSに対する良好な応答は、細胞培養条件が限定的でなかったことを示している。図18において、GGFsの細胞分裂誘発効果が、1ウェル当りに計測されたBrdU−ポジティブ及びBrdU−ネガティブ細胞の数として表されている。データは、全く同一条件で実行された2つの実験を示すものであり、少なくとも一つのウェル毎に3つのフィールドが数えられた。シュワン細胞の観察において、低投与量における増殖性効果に加えて、GGFsは、非反応性細胞の生存数も増加させる。BrdU−ポジティブ細胞の率は、前記培養に添加されたGGFsの増加量に比例する。高投与量のGGFsの存在下における48時間後の細胞の総数は、少なくとも2倍に増加し、これはGGFsがBHK21 C13細胞におけるDNA合成と増殖とを誘発することを確証するものである。同じ条件下において、2%のFCSの存在下において48時間維持した細胞は、約6倍の増加を示した(図示せず)。
【0135】
C6グリオマ細胞は、グリア細胞の特性の研究のための有用なモデルを提供した。発現した表現型は、細胞継代に依存し、細胞は、初期段階においては星状膠細胞表現型に類似し、後期段階(継代数70以降)においては乏突起膠細胞表現型に類似するようである。これらの実験において使用されたC6細胞は、継代数39から継代数52からのものであった。C6細胞はきわめて増殖性の高い個体群であるので、BrdU組み込みのバックグランドを非常に低くするように実験条件を最適化した。FCSに対する投与量反応に示されるように、細胞分裂誘発反応に大きく影響することなく細胞の生存力を維持するのに、0.1%の血清の存在が必要であった(図19)。
【0136】
図20において、aFGF(酸性線維芽細胞成長因子)とGGFsとに対する細胞分裂誘発反応が、FCS(8%)の存在下において得られた最大のBrdU組み込みの百分率として表されている。値は、全く同一の条件で実行された二つの実験の平均値である。GGFの効果は、aFGFの純粋調合物の効果と等価である。C6細胞の独特の成長因子としてaFGFが既に記載されており(Lim R et al.,1:741−746,1990)、その理由で、これはポジティブコントロールとして使用された。BrdUポジティブ及びネガティブ細胞を直接に数えることは、マイクロ培養の細胞濃度が高かったために不可能であった。これまでに報告された細胞系とは対照的に、PC12細胞は、このPC12が血清に対して反応可能な培養条件下(細胞の維持において通常に使用されるFCSとHSとの混合物)において処理された場合GGF2に対してはっきりとした反応を示さない。しかしながら、各ウェル毎のプレート化された細胞の数は、PC12細胞の挙動に影響するようである。従って、更に実験が必要である。
【0137】
例4.GGF−I及びGGF−IIペプチドを含有するタンパク質をコード化するヌクレオチド配列の分離とクローン化
GGF−IIヌクレオチド配列の分離とクローン化とを、ペプチド配列情報とライブラリースクリーニングとを使用した結果を外観し、そして以下に記載の方法によって実際に行った。尚、図4及び5のペプチドは、下記の技術に従ってGGF−I配列を分離及びクローン化する出発点として利用することができることは評価されるであろう。即ち、図21(配列認識番号54〜88)は、この目的のために利用可能な縮重オリゴヌクレオチドプローブを示し、図23(配列認識番号90〜119)は、利用可能なPCRプライマーをリストアップしている。DNA配列とポリペプチド配列とは、GGF−IIと同様にこの手段によって入手されるべきであり、更に、このようなDNA配列を組み込んだDNA構築体と発現ベクター、このような構築体/ベクターを組み込むことによって遺伝子的に変成されたホスト細胞、また、このようなホスト細胞を培養することによって得られるタンパク質も同じである。本発明は、更に、これらの物質をも含む。
【0138】
I.オリゴヌクレオチドプローブ及びプライマーの構成と分析
前記アミノ酸配列(精製GGFタンパク質からのペプチド由来のもの)をヌクレオチド配列に逆転写(backtranslate)することによって縮重DNAオリゴマープローブを構成した。オリゴマーは、前記DNA配列のコード化ストランド又は非コード化ストランドを表した。オリゴマー構成にセリン、アルギニン又はロイシンが含まれていた場合には、曖昧さをなくすために二つの別の合成物を作った。例えば、セリンは、537及び538又は609及び610のTCN又はAGYのいずれかによってコード化された。アルギニン又はロイシン(例えば、544,545)について類似のコドン分割を行った。DNAオリゴマーを、0.2マイクロモルスケールでのβ−シアノエチル化学反応を使用してBiosearch87504−カラムDNAシンセサイザーで合成した。オリゴマーを、前記カラム(500オングストロームCpG樹脂)から切り離し、濃い水酸化アンモニウム中において55〜60℃で6〜24時間保護解除(de−protected)した。これら保護解除オリゴマーを真空下で乾燥させ(Speedvac)、7Mの尿素を含有する、15%アクリルアミド(20モノ:1ビス)50mM Tris−ホウ酸−EDTAバッファのゲル中での電気泳動によって精製した。UVシャドウイング(shadowing)によって、前記ゲル中に全長オリゴマーが検出され、次に、前記バンドを摘出し、DNAオリゴマーを4〜16時間のシェイキングによって、1.5ml s H2Oに溶出した。この溶出液を乾燥し、0.1ml H2O中に再溶解し、光吸収測定を260nmで行った。
【0139】
濃度は次の式に基づいて測定した。
(A 260×単位/ml)(60.6/長さ=x μM)
【0140】
すべてのオリゴマーを、H2Oの添加によって、50μMの濃度に調節した。
【0141】
上記構成の縮重プローブが、図21、配列認識番号54〜88に示されている。
【0142】
下記の変更を加えたうえでプローブに対して使用したものと実質的に同じ手順によってPCRプライマーを作った。13のヌクレオチド含有制限サイトのリンカーが、ベクターへのクローン化用の前記縮重オリゴマーの5’末端に含まれていた。DNA合成を、1,000オングストロームCpG樹脂を使用した1マイクロモルスケールで行い、すべての4つのヌクレオチドが縮重プローブに正常に取り込まれた位置においてはイノシンを使用した。PCRプライマーの精製には、前記ゲル電気泳動精製後にエタノール沈澱することを含んでいた。
【0143】
II.ライブラリー構築及びスクリーニング
ウシゲノムDNAライブラリーを、Stragagene(カタログ番号:945701)から購入した。このライブラリーは、ベクターラムダダッシュIIにクローン化された、2×106の15〜20kb Sau3Al部分ウシDNA2フラグメントを含有していた。ウシの全脳CDNAライブラリーを、Clonetech(Catalogue Number:BL10139)から購入した。全脳と、ウシ下垂体と、ウシ後部下垂体とから、調製したmRNAから相補性DNAライブラリーを構成した(In Vitrogen;Stragene)。Vitrogenは2つのcDNAライブラリーからなる:一つのライブラリーは、ベクターラムダg10であり、他方はベクターpcDNAIである(プラシミドライブラリー)。前記Stratagenライブラリーは、ベクターラムダユニザップ(unizap)よりなる。全体として、前記cDNAライブラリーは、1,400万の一次組替えファージを含有していた。
【0144】
前記ウシゲノムライブラリーを、各プレートにつき、150,000ないし200,000のファージプレークで、E.coli K12ホスト株LE392で23x23cmのプレート(Nunc)にプレート化した。各プレートは、約一つのウシゲノム等価物を表した。37℃での一晩の培養後、前記プレートを冷却して、マニアティス(Maniatis)他(1:60−81)の手法に従って、複製フィルターを作った。4つのプレークリフトを、各プレートから非荷電ナイロン膜に作った(Pall Biodyne A又はMSI Nitropure)。前記DNAを、紫外線下で5分間のクロスリンキングによって、又は、80℃の真空下において2時間のベーキングで、前記膜上に固定した。DNAプローブを、製造会社の指示に従って、ガンマ32P ATP(New England Nuclear;6500 Ci/mmol)とともにT4ポリヌクレオチドキナーゼ(New England Biolabs)を使用して標識化した。簡単に説明すると、50pmolsの縮重DNAオリゴマーを、37℃で30分間、600μCi ガンマ32P−ATPと5単位のT4ポリヌクレオチドキナーゼの存在下において培養した。反応を終結させ、ゲル電気泳動負荷バッファを添加し、次に、放射能標識化プローブを、電気泳動によって精製した。
【0145】
32P標識化プローブを、ゲルスライスから摘出し、水に溶出した。別の方法として、DNAプローブを、ショウォルター(Schowalter)及びソマー(Sommer),Anal.Biocem177:90〜94(1989)の実験記録に従って、α−32P−dATP又はα−32P dCTPを組み込むことにより、PCR増幅で標識化した。PCR反応において標識化したプローブを、セファーデックスG−150カラムでの脱塩によって精製した。
【0146】
プレハイブリダイゼーションとハイブリダイゼーションを、GMCバッファ(0.52M NaPi,7% SDS,1% BSA,1.5mM EDTA,0.1N NaCl 10mg/ml tRNA)中で行った。洗浄は、オリゴウォシュ(160ml 1MNa2HPO4,200ml 20% SDS,8.0ml 0.5M EDTA,100ml 5M NaCl,3632ml H2O)。通常は、10のウシゲノム等価物の複写コピーを表す20のフィルター(それぞれ400平方センチ)を、100pmolsの縮重オリゴヌクレオチドプローブ(128−512倍の縮重)とともに200mlのハイブリダイゼーション溶液中において培養した。ハイブリダイゼーションは、前記縮重プローブに対し算出された最低融解温度以下の5℃で、一晩、進行させるようにした。尚、最低融解温度の計算においては、ATペアに対して2℃、GCペアに対して4℃と仮定した。
【0147】
フィルターを、そのハイブリダイゼーション温度において、オリゴウォシュを繰り返し交換して、4ないし5時間にわたり洗浄し、最後に、DNAプローブの長さに依存した温度において30分間、3.2Mテトラメチルアンモニウムクロライド、1%SDSで2度洗浄した。20mersの場合、最終洗浄温度は60℃であった。フィルターを取り付け、次に、強化スクリーン(デュポン社、Cronex Lightening Plus)を使用してX線フィルム(コダック社XAR5)に露光した。通常は、−80℃において3ないし5日間の露光で、これらのライブラリースクリーン中の複製信号を検出するのに十分であった。その結果の分析後、フィルターを取り外して、再プローブすることができた。フィルターの取り外しは、10mMのEDTA pH8を含有する1%SDSの溶液中で、フルパワー状態のマイクロウェーブオーブンで15分間のサイクルを2回行って培養することによって行われた。フィルターは、最低3ないし4回のサイクルを通じて取られ、様々なプローブで再プローブした。
【0148】
III.組替えファージの分離、成長及びDNA調製
これらの手法は、組替えDNA(マニアティス(Maniatis)他2:60−2:81)に記載の標準実験記録に基づいて行われた。
【0149】
IV.DNA消化とサザンブロットとを使用した分離クローンの分析
組替えファージDNAサンプル(2マイクログラム)を、制限エンドヌクレアーゼ製造会社(New England Biolabs)によって推奨された条件に従って消化した。37℃に於ける4時間の培養後、反応生成物を、0.1M酢酸ナトリウムと3倍量のエタノールとの存在下において沈澱させた。沈澱DNAを、遠心分離によって採取し、75%のエタノールですすぎ、乾燥させた。すべての再懸濁サンプルを、アガロースゲル(通常、TAEバッファにおいて1%0.04M Trisアセテート、0.002M EDTA)にかけた。ゲルのランは、1センチメートル当り1ボルトで4ないし20時間行った。マーカーには、ラムダHind III DNAフラグメント及び/又はφX174HaeIII DNAフラグメント(New England Biolabs)が含まれていた。前記ゲルを、0.5マイクログラム/mlの臭化エチディウムによって染色し、撮影した。サザンブロッティングのために、DNAを、先ず0.125N HClでの処理によって、前記ゲル中にて脱プリンし、0.5N NaOHで変成し、そして20×SSC(3M 塩化ナトリウム、0.03M クエン酸塩ナトリウム)で非荷電ナイロン膜に移した。ブロッティングを、6時間から24時間かけて行い、次に、フィルターを、0.5Tris HCl pH7.5,0.15M塩化ナトリウム中で中和し、次に、50mMのTris−ホウ酸 EDTA中で簡単にすすいだ。
【0150】
架橋するために、前記フィルターを、先ず、透明プラスチックラップに包み、次に、DNA側を紫外線に5分間露光した。ハイブリダイゼーションと洗浄を、ライブラリースクリーニングに関して説明したように行った(この実施例の第2節参照)。類似の遺伝子が他の種に存在するか否かを調べるためのハイブリダイゼーション分析において、僅かな変更を行った。前記DNAフィルターは、Clonetech(Catalogue Number7753−1)から購入したもので、各レーン当り様々な種に関しての5マイクログラムのEcoRI消化DNAを含有している。前記プローブを、前記セクション2に記載の方法で、PCR増幅反応によって標識化し、ハイブリダイゼーションを、硫酸デキストランを10%を含有する80%バッファB(2gのポリビニルピロロリジン、2gのFicoII−400、2gのウシ血清アルブミン、50ml 1M Tris−HCl(pH7.5)58g NaCl,1gピロ燐酸ナトリウム、10gのドデシル硫酸ナトリウム、950mlの水)中で行った。前記プローブを、10分間の沸騰で変成させ、次に、氷水中にて急冷させた。このプローブを、ミリリットル当り106dpm32Pで前記ハイブリダイゼーションバッファに添加し、60℃で一晩培養した。前記フィルターを60℃にて、先ずバッファB中で、次に2XSSC、0.1%SDSで、その後、1xSSC,0.1%SDSで洗浄した。ストリンジェンシーを高くするために、実験、最終洗浄は、0.1xSSC、1%SDSで、温度を65℃に上げて行った。
【0151】
前記ゲノムクローンの制限マップを作り、どのサブフラグメントが前記GGFプローブ(サブクローニングの候補)にハイブリダイゼーションしたかを示すためにサザンブロットを使用した。
【0152】
V.DNA相同体のセグメントのハイブリダイゼーションプローブへのサブクローニング
DNA消化物(例えば、5マイクログラム)を、1%のアガロースゲルに投入し、次に、適当なフラグメントを染色後、ゲルから摘出した。前記DNAを、ガラスビーズへの吸着によって精製し、その後、製造会社(Bio 101)によって記載された実験記録を使用して溶出した。回収したDNAフラグメント(00〜200ng)を、T4リガーゼ(New England Biolabs)を使用して、例えば、pUC18の派生物であるpT3T7(Ambion)等のリニア化された脱燐酸化ベクター中に結合した。このベクターは、E.coli.βラクタマーゼ遺伝子を有しているので、形質転換物質を、アンピシリンを含むプレート上で選別することが可能である。前記ベクターは、更に、ホスト細胞へβ−グラクトシダーゼ相補性を与えるので、非−組替え(ブルー)を、イソプロピルチオガラクトシドとブルオーグ(Bluog)他(Bethesda Research Labs)とを使用して検出することができる。前記結合(ligation)反応物の一部を、使用して、E.coli K12 XL1ブル競合細胞(Stratagene Catalogue Number 200236)を形質転換し、次に、これらの形質転換物質をアンピシリン1ml当り50マイクログラムを含有するLBプレート上にて選別した。ホワイトコロニーを選別し、プラスミドミニプレップをDNA消化及びDNA配列分析のために調製した。選別したクローンを、それらの挿入DNAが前記GGFプローブとハイブリダイズしたか否かを確認するために再テストした。
【0153】
VI.DNA配列決定
2重ストランドプラスミドDNAテンプレートを、標準実験記録に従って、5mlの培養から調製した。製造業者の実験記録に従い、シーケナーゼ2.0とジデオキシヌクレオチド配列キット(US Biochemical)とを使用したジデオキシチエーン終結法によって、配列決定を行った(サンガー(Sanger)他PNAS;USA74:5463(1977)の変形)。あるいは、配列決定を、サイクルシークエンシングキット(New England Biolabs;Bethesda Research Laboratories)を使用したサーマルサイクラー(Perkin Elmer,model 4800)中において行い、5’−末端標識化プライマーを使用した製造業者の指示に従って行った。配列プライマーは、前記配列決定キットで供給されたものか、あるいは、前記クローンから決められた配列によって合成されたものであった。配列決定用反応物を、6%ポリアクリルアミドの0.4mm厚さ配列決定用ゲル上に負荷(load)し、溶解した。ゲルを乾燥し、X線フィルムに露光した。通常、標準配列決定キットが使用された時は35Sが組み込まれ、32P末端標識化プライマーをサイクルシークエンシング配列決定反応に使用した。配列は、ゲルの底部から頂部の方向(5’より3’の方向に向かって)にDNA配列エディタに読み込まれ、データを、ジェネティクス・コンピューター・グループ(Genetics Computer Group)(GCG,ウィスコンシン大学)によって供給されているプログラムを使用して解析した。
【0154】
VII.RNAの調製とPCRの増幅
ゲノムDNA中に検出され、GGFペプチドをコード化する配列を含む転写読み取り枠を、下垂体RNAのPCR増幅によって拡張した。RNAは、グアニジン中性−CsCl手法(チャーグウィン(Chirgwin)他Biochemistry18:5294(1979))に従って冷凍ウシ繊維組織(Pelfreeze)から調製した。ポリアデニール化RNAは、オリゴ−dTセルロースカラムクロマトグラフィによって選別した(Aviv and Leder PNAS(USA)69:1408(1972))。
【0155】
特定のDNA目標配列を、全RNA、もしくは、Perkin Elmer PCR/RNAキットNo.N8−8−0017を使用してcDNAから変換しておいたポリアデニール化RNAサンプルから始めて増幅した。第1ストランド逆転写反応は、1μgのテンプレートRNA及び、制限酵素認識サイトリンカーを結合したオリゴdTのプライマーカー、又は、制限サイトを結合したクローン化配列から決められた特定のアンチセンスプライマーのいずれかを使用した。第2ストランドを作るために、前記プライマーは、3’RACE反応(フローマン(Frohman)他PNAS(USA)85:8998(1988))において使用されるプラスストランド特徴配列か、あるいは、第2目標サイトが、予め、dATPを備えた第1ストランド反応生成物の後の終端トランスフェラーゼによって添加された場合(例えば、5’のレース反応、フローマン(Frohman)他ibid)には制限サイトを取り付けたオリゴdTプライマーのいずれかであった。あるいは、固定されたPCR反応物と同様に、前記第2ストランドプライマーは分解物であって、従って特定のペプチド配列を示すものであった。
【0156】
増幅プロフィールは、次の一般的手法に従った。
1)95℃で5分間の浸漬ファイル;2)95℃、1分間の熱サイクルファイル、1分間で45℃、50℃又は55℃のアニーリング温度にまで下げ、このアニーリング温度を1分間維持:1分間以上で72℃にまで温度を上げる;72℃で伸長するか、あるいは、1分10秒で自動伸長、3)72℃での5分間の伸長サイクル、及び4)4℃で不定時間の浸漬ファイル。熱サイクルファイル(#2)は、通常、30サイクルだけ実行した。各100μlの増幅反応の16μlサンプルを3時間、1センチメータ当り4ボルトの条件で、TAEバッファ中において2%Nusieve、1%アガロースゲルでの電気泳動実行によって分析した。これらゲルを染色し、次に、前記プライマーの内部の標識化DNAプローブでプローブした非荷電ナイロン膜にブロットした。
【0157】
このブロッティング実験において、特定のセットのDNA増幅生成物を同定することができた。そして、その位置を、精製と再増幅のためのガイドとして使用した。適当な場合、選別されたサンプルの残りの部分を、調製ゲルに負荷し、次に、電気泳動を行い、前記ゲルから0.5mm厚さの4ないし5のスライス(特定生成物の予想位置をブラケット化する)を取り出した。前記アガロースを粉砕し、次に、40℃で2〜16時間、0.5mlの電気泳動バッファ中に浸漬した。粉砕したアガロースを2分間遠心分離し、水相を新しい試験管に移した。
【0158】
最初の反応と同じセットのプライマーと反応プロフィールとを使用し、5μl(生成物の約1%)の溶出物質に対して再増幅を行った。再増幅反応が完了したとき、サンプルを、クロロフォルムで抽出し、新しい試験管に移した。リンカー中に存在する制限サイトで切り離すために、濃縮制限酵素バッファと酵素とを、反応物に添加した。消化PCR生成物を、ゲル電気泳動によって精製し、次に、前述のサブクローン化の章に記載のようにベクターにサブクローン化した。DNA配列決定は、前述のように行った。
【0159】
VIII.DNA配列分析
フラグメントアセンブリプログラムを使用してDNA配列をアセンブルし、アミノ酸配列を、GCGプログラムGel Assemble,Map and Translateによって推定した。これらの推定タンパク質配列を、質問配列(query sequence)として使用して、ワード・サーチ(Word Search)を使用してタンパク質配列データベースを検索した。分析は、VMS5.1で作動するVAXステーション3100ワークステーションにて行った。前記データベース検索は、GCGバージョン7.0を使用したSwiss Prot release No.21にて行った。
【0160】
IX.GGF−I及びGGF−IIをコード化する遺伝子のクローン化と配列決定の結果
前述のように、ウシGGF−IIをコード化するDNA配列を同定するために、分解オリゴヌクレオチドプローブを、GGF−IIペプチド配列から構成した。リシルエンドペプチダーゼによって消化された、精製GGF−II調合物のペプチドであるGGF−II12(配列認識番号44)(図11及び12参照)は、精製GGF−I調製物から調製されたトリプシンペプチドであるGGF−I 07(配列認識番号39)と強いアミノ酸配列相同性を示した。従って、10個の縮重オリゴヌクレオチドプローブを作るためにGGF−1112を使用した(図21の配列認識番号69,70,71,79としてそれぞれ示されているオリゴ609,610,649〜656参照)。フィルターの複製セットを、GGF−II 12の二つのオーバーラップする部分をコード化する2セットのプローブ(セット1=609,セット2=649〜5656)でプローブした。ハイブリダイゼーション信号が観察されたが、両方のプローブセットにハイブリダイズしたクローンは一つだけであった。このクローン(GGF2BG1として指定)を精製した。
【0161】
前記ファージクローンGGF2BG1からのDNAのサザンブロッティング分析によって、両方のプローブセットが前記ウシDNA配列とハイブリダイズしたことが確認されるとともに、更に、両方のプローブが、該クローン中において、同じセットのDNAフラグメントと反応していることが判った。これらの実験に基づき、元のクローンの4kb Eco RIサブ−フラグメントが同定され、サブクローン化され、部分的に配列決定された。図22は、前記ヌクレオチド配列認識番号89とプローブ609及び650のハイブリダイゼーションサイトを含む初期DNA配列解読の推定アミノ酸配列を示し、更に、このウシゲノムDNAの部分がペプチド12(KASLADSGEYM)をコード化したことを立証している。
【0162】
更に配列分析をしたところ、前記GGF−II 12が、ウシGGF−II遺伝子とcDNAとを表すオーバーラップ配列の分離の開始点になった66アミノ酸転写読み取り枠(下記参照)に位置することが判った。
【0163】
いくつかのPCR手法を使用して、推定ウシGGF−II遺伝子の、更に別のコード化配列を得た。全部のRNA及びオリゴdT−選択(ポリA含有)RNAサンプルを、ウシの全下垂体、前方下垂体、後方下垂体、及び視床下部から調製した。図23、配列認識番号109〜119に示すリストからのプライマーを使用して、片側(one−sided)PCR反応によって、3’と5’との両方の方向においてcDNA末端を増幅し、固定PCR反応を、別のGGF−IIペプチドを表す縮重オリゴヌクレオチドプライマーにて行った。図24は、これらの実験において得られた隣接するDNA構造体と、配列とをまとめて示している。3’レース(RACE)反応から、三つの交互にスプライスされたcDNA配列が生成され、これらをクローン化し配列決定した。5’レース反応によって、少なくとも52のアミノ酸のコード化配列を含む別のエクソンが発見された。この推定アミノ酸配列の分析によって、ペプチドGGF−II−6と、GGF−I−18に類似した配列とが明らかになった(下記参照)。固定化PCR反応によって、300bpの別のcDNAセグメント中に含有されるペプチドGGF−II−1,2,3及び10の(cDNA)コード化配列が同定された。このセグメントの5’境界(即ち、図31のセグメントE)は、ペプチドGGF−II−1をコード化するとともに、前記PCR反応において使用されたオリゴヌクレオチドによって規定されている(例6のヒトクローンに関して記載されているように別の5’配列データが存在する)。従って、このクローンは、既存の全部で9つの新規なGGF−IIペプチド配列の内の6つをコード化するヌクレオチド配列を有する。
【0164】
前記クローン化遺伝子は、先ず、前記コード化配列を、それらが見出された状態(図25参照)において位置決めすることを可能にするGCF2BG1の物理的地図を構成することによって特徴付けられる。上述のコード化配列からのDNAプローブを使用して、このファージクローンにおいて前記エクソンを含有する更に別のDNAフラグメントを同定するとともに、両方の方向においてオーバラップするクローンを同定した。前記推定ウシGGF−II遺伝子は、少なくとも5つのコード化セグメントに分割される。コード化セグメントは、ユニバーサルな遺伝子コードを使用してポリペプチド配列に翻訳可能な、個々の長さのDNA配列であると定義される。図31に示し、本出願において言及されるこれらコード化セグメントは以下の通りである。1)GGF遺伝子内に存在する特定のエクソン(例えば、コード化セグメントa)、あるいは、2)各セットが図示の遺伝子生成物と同様に特定のポリペプチドセグメントに翻訳可能な場合において、mRNAsの特定のサブーグループ中に現れる2つ又はそれ以上のエクソンのセットから派生するもの。クレームにおいて言及されているポリペプチドセグメントは、前記相同DNAコード化セグメントの翻訳生成物である。これまで、コード化セグメントA及びBのみが、エクソンとして定義され、配列決定されマップ化された。図26は、同定された隣接コード化配列をまとめて示している。エクソンは、その発見の順序(アルファベット順)でリストされている。そのイントロン/エクソン境界から、エクソンBを、コード化セグメントEとコード化セグメントAとを接続するcDNAsに含めることが出来ることが明らかである。即ち、エクソンBは、その読み取り枠を損なうことなく、スプライスすることは出来ない。従って、我々は、三つのスプライシングパターンが、推定ウシGGF−II cDNA配列1,2及び3を生成することが出来ると示唆する。それぞれ、GGF2BPP1.CDS、GGF2BPP2.CDS及びGGF2BPP3.CDSとされるこれらのコード化配列は、図28A(配列認識番号133)、28B(配列認識番号134)及び28C(配列認識番号135)にそれぞれ示されている。これら三つのcDNAの推定アミノ酸配列も、図28A(配列認識番号133)、28B(配列認識番号134)及び28C(配列認識番号135)にそれぞれ示されている。
【0165】
これら三つの推定構造体は、アミノ酸長さが206,281,257であるタンパク質をコード化する。推定されたタンパク質配列の最初の183残基は3つの遺伝子生成物において同一である。位置184において、前記クローンは大幅に異なっている。GGF2BPP1におけるグリシンGGT用に対するコドンは、又、GGF2BPP2とGGF2BPP3に対するスプライスドナーとしても作用し、これらは、それぞれ交互に、エクソンC,C/D,C/D’及びD又はC,C/D及びDに加わるものであり、図33において、配列認識番号149として示されている。GGFIIBPP1は、前記コード化セグメントAスプライス接合部を越えて次の介在配列(イントロン)への読み取りによって生成される裁頭遺伝子生成物である。これは、図31におけるコード化セグメントA’を示す(配列認識番号140)。前記転写体は、標準型AATAAAポリアデニール化配列の隣で終端し、我々は、この裁頭遺伝子生成物が、真性成熟転写体を表すものであると提案する。他の二つのより長い遺伝子生成物は、同じ3’非翻訳配列とポリアデニリン化配列を共有する。
【0166】
これらの三つの分子は、すべて、9つの新規なGGF−IIペプチド配列の内の6つを有し(図12参照)、もう一つのペプチドがGGF−I−18に対してきわめて高い相同性を有している(図27参照)。この発見は、この組替え分子が、ウシGGF−IIの少なくとも一部分をコード化している可能性がきわめて高いことを示している。更に、三つのペプチドの計算上の等電点は、GGF−IとIIの物理的特性と一致している。GGF−IIの分子の大きさはおよそ60kDであるので、これら三つのcDNAの最も長いものが、予想数のアミノ酸の1/2近くをコード化しているはずである。
【0167】
B及びAエクソンを含むプローブを、PCR増幅によって標識化し、ウシ後部下垂体から分離されたRNAから作られたcDNAライブラリーのスクリーニングに使用した。一つのクローン(GGF2BPP5)は、図30に示すパターンを示し、コード化セグメントAとCとの間にもう一つのDNAコード化セグメント(G)を有していた。核酸配列の全体が図32に示されている(配列認識番号148)。最長転写読み取り枠からの予想翻訳生成物は、241のアミノ酸である。第2のcDNAの一部(GGF2BPP4)も、上述のプローブを使用してウシ後部下垂体ライブラリーから分離された。このクローンは、図30に示すパターンを示した。このクローンは、5’の部分において不完全であるが、コード化セグメントG及びDを欠いているという意味においてこれはスプライシング変異体である。BPP4も、領域C/D以降において領域H,K及びLを備えた新規な3’末端を示す。BPP4の配列は、図34に示されている(配列認識番号150)。
【0168】
例5
様々な種に於けるGGF配列
データベース検索によっては、予想GGF翻訳生成物と公知のタンパク質配列との間になんら有義な類似性は見つからなかった。これは、GGF−IIが、タンパク質の新規なファミリ又はスーパーファミリの最初のメンバであることを示唆している。これまでに我々が示した、他のほ乳類DNAとの高ストリンジェンシーのクロスハイブリダイゼーション研究(DNAブロッティング実験)においては、はっきりと、このウシ組替え分子からのDNAプローブによって、テストされた様々なサンプルにおいて特定の配列が容易に検出される。相同性の高い配列は、ヒトゲノムDNAにも検出された。そのオートラジオグラムを図29に示す。ラットとヒトのDNAを含むレーンの信号は、GGF遺伝子のラット及びヒト同等物を表し、この遺伝子によってコード化された複数のcDNAの配列が、最近、ホルムズ(Holmes)他(Science 256:1205(1992))とウェン(Wen)他(Cell 69:559(1992))とによって報告されている。
【0169】
例6
ヒト配列コード化ヒトGGF2の分離
ウシGGFIIコード化セグメントEをからの配列を含む複数のヒトクローンを、脳幹から調製されたヒトcDNAライブラリーをスクリーニングすることによって分離した(Stratagnene catalog#935206)。このストラテジーを、大半のGGF2ペプチド(GGF2に特有)と、前記ウシEセグメントを含むクローンからの予想ペプチド配列との間の強い関連性に基づき、追求した。このライブラリーは、次にリストするオリゴヌクレオチドプローブ914−919を使用した前述の例4、第11章で記載したものと同様にスクリーニングした。
【0170】
これらのプローブで検出されたクローンを、ハイブリダイゼーションによって更に分析した。セグメントAからのポリメラーゼ連鎖反応(PCR)生成物を標識化することによって生成された、コード化セグメントAからのプローブ(図21参照)は、一次ライブラリーをスクリーニングするのにも利用した。A及びE由来のプローブとハイブリダイズした複数のクローンを選別し、一つの特定のクローン、GGF2HBS5、を選別して更に分析した。このクローンは、コード化セグメントのパターン(図31に示すEBACC/D’D)によって表されている。このクローン中のEセグメントは、図37に示されるEの裁頭化ウシ・バージョンのヒト等価物である。GGF2HBS5は、前述のすべての「推定」GGF−II候補の内で最も可能性の高い候補である。コード化配列セグメントEの長さは、786のヌクレオチドと264の非翻訳配列のベースとである。GGF2HBS5によってコード化されるタンパク質の予想サイズは、約423アミノ酸であり(約45キロダルトン、図45、配列認識番号170参照)、これはデグリコシル化状態のGGF−II(例16参照)のサイズに類似している。更に、図27にリストしたGGF−IIペプチドのうちの7つは、領域Eから予想されるタンパク質配列内に属する同等配列を有している。ペプチドII−6とII−12は例外であって、それぞれコード化セグメントB及びコード化セグメントAに属する。GF2HBS5タンパク質をコード化するRNAを、GGF2HBS5挿入部を有するベクター(Bluescript SK[Stratagene Inc.]図44参照)にあるバクテリオファージT7プロモーターによってドライブされた生体外転写システム中において生成した。このRNAは、無細胞(ラビット網状赤血球)翻訳システムに翻訳され、そのタンパク質生成物のサイズは45kdであった。
【0172】
更に、前記無細胞生成物を、シュワン細胞細胞分裂誘発分析において分離し、生物学的活性を確認した。調整培地によって処理されたシュワン細胞は、125I−ウリジンの組み込みによって測定した増殖度の増加と、185キロダイン領域のタンパク質のチロシンの燐酸化との両方を示した。従って、GGF2HBS5によってコード化された生成物のサイズと、図12に示したウシペプチドに対する相同性の高いヒトペプチドをコード化するDNA配列の存在とは、GGF2HBS5が、ウシGGF2のヒト等価物をコード化することを裏付けるものである。このクローンと形質変換した細胞から作られた調整培地が、シュワン細胞細胞分裂誘発活性を顕在化させるという事実から、GGFIIHBS5遺伝子生成物(前記BPP5遺伝子生成物とは違って)が分泌されるということが確かめられた。更に、GGFIIBPP5遺伝子生成物は、p185erbB2等のレセプターチロシンキナーゼや、あるいは、密接に関連したレセプター(図14参照)を介してシュワン細胞増殖反応を仲介すると考えられる。
【0173】
例7
ほ乳類及び昆虫細胞におけるヒト組替えGGF2の発現
ヒトGGF2をコード化する前記GGF2HBS5cDNA(例6において記載され、ここでHBS5と称される)を、ベクターpcDL−SRα296にクローン化し(タケベ(Takebe)他Mol.Cell.Biol.8:466−472(1988))、COS−7細胞を、DEAE−デクストラン法(サムブルック(Sambrook)他 Molecular Cloning:A Laboratory Manual 2nd ed.CSH Laboratory NY(1989))によって、100mmディシュにおいて移入させた。細胞溶解物又は一過的発現COS細胞からの調整培地を、移入の3又は4日後に収穫した。溶解物を調製するために、細胞単層を、PBSによって洗浄し、150μlの0.25M Tris−HCl,pH8中での三回の冷凍/解凍サイクルによって溶解したディシュから掻き取った。細胞堆積物をペレットで取り、上澄を回収した。調整培地サンプル(7ml)を収集し、次に、濃縮し、製造業者(アミコン(Amicon),マサチューセッツ州,ビヴァリー)によって記載されたようにCentiprep−10とCentricon−10ユニットを使用して、10mM Tris,pH7.4とバッファ交換した。前述したように(例3参照)、ラットの神経シュワン細胞のDNA合成先駆体の組み込みを分析した。調整培地又は細胞溶解物サンプルを、例3において記載した方法で、シュワン細胞増殖分析においてテストした。その細胞分裂誘発活性データは、図46に示されている。GGF2をコード化する、cDNA、GGF2HBS5は、タンパク質生成物の分泌を前記培地に振り向けた。全活性の極く一部分が、細胞分解物を使用した分析によって、細胞内に検出可能であった。GGF2HFB1とGGFBPP5 cDNAでは、生成物の分泌を細胞外培地に振り向けることが出来なかった。これらのクローンからのGGF活性物質は、細胞分解物においてのみ検出可能であった(図46)。
【0174】
組替えGGF2も、CHO細胞に発現された。GGF2をコード化するGGF2HBS5 cDNAを、ベクターpcdhfrpolyAのEcoRIサイトにクローン化し(図54)、燐酸カルシウム共沈法(Graham and Van Der Eb,Virology 52:456−467(1973))によってDHFRネガティブCHO細胞に移入した。クローンを、96−ウェルプレート中のヌクレオチド及び無ヌクレオチドα培地(Gibco)内で選別した。3週間後、個々のクローンの調整培地を、例3で記載したシュワン細胞増殖分析によって、GGF発現のためにスクリーニングした。前記培地へかなり高いレベルのGGF活性物質を分泌した安定したクローンが同定された。CHO細胞調整培地の異なった分量の部分標本から得られたシュワン細胞増殖活性データを使用して、図47に示す投与量反応曲線を得た(Graham and Van Der Eb,Virology 52:456,1973))。この物質を、GGF2特定ペプチドに対して生成したポリクローナル抗血清によってプローブしたウェスタンブロッティングにて分析した。約69〜90kd(下垂体及び高分子量グリコフォームから抽出されたGGF2の予想サイズ)の広い範囲のバンドが特異的に標識化されている(図49,レーン12)。
【0175】
組替えGGF2は、バキュロウィルス発現を使用して昆虫細胞にも発現した。Sf昆虫細胞を、GGF2HBS5cDNAクローンを含有するバキュロウィルスに、3〜5の多重度(106細胞/ml)で感染させ、そしてSf900−II培地(Gibco)中で培養した。シュワン細胞細胞分裂誘発活性物質を、前記細胞外に培地中に分泌させた(図48)。異なった分量の昆虫細胞調整培地を、フォルスコリン不存在の状態で、シュワン細胞増殖分析でテストし、そのデータを使用して図48に示す投与量反応曲線を得た。
【0176】
この物質は、更に、前述のGGF II特異性抗体でプローブしたウェスタンブロッティング(図47)でも分析した。45kdのバンド、デグリコシル化GGF−II(例16)のサイズが示されている。
【0177】
この例で使用された方法は、次の通りである。
【0178】
組替えヒト及びウシ・グリア成長因子のシュワン細胞細胞分裂誘発活性を以下のように測定した。培養シュワン細胞の細胞分裂誘発反応を、短命ほ乳類発現実験から得た粗組替えGGF生成物を使用して、5・lのフォルスコリンの存在下において測定した。[125I]−ウリジンの組み込みを、前述の方法に記載の移入した、あるいは疑似移入したCOS細胞から得た物質に対して18〜24時間さらした後に行った。4セットのデータの平均及び標準偏差が図示されている。部分的に精製された天然ウシ下垂体GGF(カルボキシルメチルセルロースフラクション:グッドアール(Goodearl)他、提出)に対する細胞分裂誘発反応が、100パーセントの活性の標準として図示(GGF)されている。
【0179】
cDNA(図53)を、pcDL−SR痰Q96にクローン化し(タケベ(Takebe)他,Mol.Cell biol.8:466〜472(1988))、COS−7細胞を、DEAE−デキストラン法(サムブルック(Sambrook)他,In Molecular Cloning.A Laboratory Manual,2nd.ed.(Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY,1989))によって100mmディシュ中で移入した。細胞溶解物又は調整培地を転写の3又は4日後に収穫した。溶解物を調製するために、細胞単層をPBSによって洗浄し、ディシュから掻き取り、150μlの0.25M Tris−HCl,pH8中での3回の冷凍/解凍サイクルによって溶解した。細胞堆積物をペレットとし、上澄を回収した。調整培地サンプル(7mls)を採取し、次に、濃縮し、製造業者(アミコン(Amicon),マサチューセッツ州,ビヴァリー)によって記載されたようにCentiprep−10とCentricon−10ユニットを使用して、10mM Tris,pH7.4とバッファ交換した。前述したように(デイヴィス(Davis)およびストローバント(Stroobant),J.Cell Biol.110:1353−1360(1990);ブロックス(Brockes)他,Brain Res.165:105〜118(1979))、ラットの座骨神経シュワン細胞はDNA合成先駆体の組み込みについて分析した。
【0180】
組替えCHO細胞調整培地のウェスタンブロッティングは、以下のように行った。一つのCHOクローンを、7mlのMCDB302無タンパク質培地中で3日間、培養した。2mlの調整培地を濃縮し、10mMのTris−HCl pH7.4に対してバッファ交換し、凍結乾燥させた。そのペレットをSDS−PAGEサンプルバッファ中で再懸濁し、還元性SDSゲル電気泳動にかけ、GGFペプチド抗体でのウェスタンブロッティングによって分析した。CHOコントロールを、非移入CHO−DG44ホストからの調整培地を使用して行い、CHO HBS5レベルを、組替えクローンからの調整培地を使用して分析した。
【0181】
例8
ウシGGFに関連する他のヒト配列の分離
例5及び6の結果は、ヒトのソースからのGGF関連配列が、ウシGGF配列由来のDNAプローブを使用することによっても容易に分離可能であることを示している。これに代えて、ホルムズ(Holmes)他(Science 256:1205(1992))によって記載された方法も利用可能である。この例において、前記p185erbB2レセプターに結合し、これを活性化する(そしてGGFに関係する)ヒトタンパク質(ヘレグリン瘁jが、腫瘍細胞系から精製され、派生したペプチド配列を使用して、cDNAコード化ヘレグリンをクローン化するのに使用されたオリゴヌクレオチドプローブを生成する。p185erbB2レセプター活性の生化学的分析は、シュワン細胞の増幅とは異なる。これは、下垂体cDNAからGGF配列をクローン化するために例1〜4において使用した方法に類似している。ヘレグリンタンパク質と相補性DNAとは、次の手法に従って腫瘍細胞系から分離した。
【0182】
ヘレグリンは、Percell Biolyticaマイクロキャリアビーズ(Hyclone Labs)上で成長したMDA−MB−231乳ガン細胞(ATCC #HTB26)によって調整した培地から精製した。前記培地(10リットル)は、膜(10−kDカットオフ)(Millipore)によるフィルター処理によって〜25倍に濃縮し、遠心分離処理と、フィルター(0.22um)を通す濾過処理とによって清浄化した。濾過物を、ヘパリンセファロースカラム(Pharmacia)にかけ、そのタンパク質を、燐酸−緩衝食塩水中にて0.3、0.6及び0.9MのNaClステップで溶出した。種々のクロマトグラフィック・フラクション中の活性を、MCF−7乳ガン細胞(ATCC#HTB22)中におけるp185erbB2のチロシン燐酸化の増加を定量化することによって測定した。MCF−7細胞を、血清(10%)を含有した(ウェル当り105の細胞)、F12(50%)のダルベッコの最小必須培地(50%)中にて24−ウェルCostarプレートにプレート化し、少なくとも24時間付着させた。分析の前に、細胞を、少なくとも1時間、血清を含まない培地中に移した。カラムフラクション(10ないし100μl)を、37℃で30分間、培養した。次に上澄を吸引して、SPD−PAGEサンプルバッファ(100μl)を添加することによって反応を終わらせた。サンプルを100℃で5分間加熱し、一部分(10ないし15μl)を、tris−グリシンゲル(4ないし20%)(Novex)にかけた。電気泳動後、タンパク質を、ポリフッ化ビニリデン(PVDF)膜上にエレクトロブロッティングし、次に、ツイーン(Tween)−20を含有する(0.05%)tris−緩衝食塩水(TBST)中にてウシ血清アルブミン(5%)でブロックした。ブロットを、ホスホチロシン(Upstate Biotechnology)に対するモノクローナル抗体(1:1000希釈)で、室温で最低1時間、プローブした。ブロットをTBSTで洗浄し、室温で最低30分間、アルカリフォスファターゼと結合したマウスの免疫グロブリンGに対する抗体(Promega)(1:7500に希釈)でプローブした。反応性バンドは、5−ブロモ−4−クロロ−3−インドイル−1−フォスフェートとニトロ−ブルーテトラゾリウムによって可視化した。免疫ブロットを、Scan Jet Plus(ヒューレットパーカード)濃度計によって走査した。非刺激MCF−7細胞の信号強度は、20ないし30ユニットであった。完全に刺激したp185erbB2は、180ないし200ユニットであった。活性のほとんどを有する0.6M NaClプールを、エタノール(30%)を含有する17mMの燐酸ナトリウム(pH6.8)中で平衡されたポリアスパルテン酸(PolyLC)カラムに適用した。前記平衡バッファ中で、0.3から0.6M NaClまでのリニアグラジエント使用して、結合タンパク質を溶出した。活性のピーク(0.45M NaClに於ける)を、TFA(0.1%)とアセトニトリル(15%)とを含有するバッファで平衡させたC4逆相カラム(SynChropack RP−4)にて更にフラクション化した。このカラムから、25から40%のアセトニトリル・グラジエント範囲にて、60分間にわたり、タンパク質が溶出した。フラクション(1ml)を採取し、その活性を分析し、tris−グリシンゲル(4〜20%,Novex)上でSDS−PAGEによって分析した。HPLC−精製したHRG−αを、SDS(0.1%)、10mMジチオトレイトール、0.1M NH4HCO3(pH8.0)中で、リシンCにて、37℃で20時間に渡って消化させ、その結果得られたフラメントを、Synchrom C4カラム(4000Å,0.2by10cm)上で溶解させた。このカラムを、0.1%のTFA中で平衡させ、次いで0.1%TFA中で1−プロパノール使用グラジエントで溶出した(ダブリュ・ジェイ・ヘンゼル(W.J.Henzel),ジェイ・ティ・スタルツ(J.T.Stults),シー・スー(C.Hsu),ディ・ダブリュ・アシュワド(D.W.Aswad),J.Biol.Chem.264,15905(1989))。クロマトログラフ・ランのピークを真空下において乾燥し、配列決定した。これらのペプチドの一つ(〜24%1−プロパノールで溶出)の配列は、[A]AEKEKTF[C]VNGGEXFMVKDLXNP(配列認識番号162)であった。括弧内の残基は、不確定であり、Xは、アミノ酸の同定が不可能であったサイクルを示している。最初の収量は、8.5pmolであり、その配列はいずれの公知のタンパク質にも対応していなかった。残基1,9,15及び22は、後に、cDNA配列中においてシステインとして同定された。これまで過負荷され(over−loaded)、そしてPVDFにブロットされたゲルから得られた〜45kDバンドの直接的配列決定は、初期収量が極めて低い(0.2pmol)、低量(lowabundance)の配列、XEXKE[G][R]GK[G]K[G]KKKEXGXG[K](配列認識番号163)を示した。これは、ヘレグリン−α(図31)のアミノ酸残基2ないし22に対応し、セリン2がproHRG−αのNH2末端であることを示唆している。NH2末端はブロックされているが、時として小量の自然にブロックされたタンパク質は、翻訳後に変成されないこともあることが観察された。前記NH2末端指定は、臭化シアンとの消化後のタンパク質のマススペクトル分析によって確かめられた。前記分離タンパク質のCOOH−末端は、はっきりとは同定されていなかったが、タンパク質分解消化物の混合配列決定によって、成熟した配列は、残基241を越えて拡張しないようである。アミノ酸残基の略称は以下の通りである。A,Ala;C,Cys;D,Asp;E,Glu;F,Phe;G,Gly;H,His;I,Ile;K,Lys;L,Leu;M,Met;N,Asn;P,Pro;Q,Gln;R,Arg;S,Ser;T,Thr;V,Val;W,Trp;そしてY,Tyr。
【0183】
cDNAクローンの源として、オリゴ(dT)−開始λgt10(ティ・ヴィ・ハイン(T.V.Huynn),アール・エイ・ヤング(R.A.Young),アール・ダブリュ・デイヴィス(R.W.Davis),λgt10及びλgt11DNAクローン化技術:A Practical Approach,D.Glover,Ed.(IRC Press,Oxford,(1984))cDNAライブラリーを、MDA−MB−231細胞からの、mRNA精製(ジェイ・エム・クルウィン(J.M.Chrwin),エイ・イー・プルズビラ(A.E.Przbyla),アール・ジェイ・マクドーランド(R.J.MacDoland),ダブリュ・ジェイ・ラッター(W.J.Rutter),Biochemistry18,5294(1979)によって構成した(ユー・ガブラー(U.Gubler)およびビー・ジェイ・ホフマン(B.J.Hoffman),Gene 25,263(1983))。13−アミノ酸配列AEKEKTFCVNGGE(配列識別番号164)(13)をコード化する下記の8重の縮重アンチセンスデオキシオリゴヌクレオチドを、ヒトコドン周波数最適値(frequency optima)(アール・ラテ(R.Lathe),J.Mol.Biol.183,1(1985))のベースに構成し、化学的に合成した。即ち、5’−CTCGCC(G OR T)CC(A OR G)TTAC(A OR G)CAGAAGGTCTTCTCCTTCTCAGC−3’(配列識別番号165)。プローブ構成の目的のため、システインを前記アミノ酸配列中の未知の残基に指定した。このプローブを燐酸化により標識化し、低−ストリンジェンシー条件下におけるcDNAライブラリーへのハイブリダイズした。前記proHRG−α タンパク質を、このライブラリー中において同定した。HRB−B1 cDNAは、proHRG−αの5’と3’との両方の末端からの配列とともに、MDA−MB−231細胞mRNAから作られた第2オリゴ(dT)−開始 λgt10ライブラリーのプローブ化によって同定された。クローン13(図2A)は、プライム化(5’−CCTCGCTCCTTCTTCTTGCCCTTC−3’プライマー(配列識別番号166);proHRG−α アンチセンスヌクレオチド33ないし56)MDA−MB−231 λgt10ライブラリーを5’HRG−α 配列によってスクリーニングすることによって得られた生成物であった。前記プローブとしてのクローン13の5’末端に対応する配列を使用してMDA−MB−231細胞mRNAからの第3のオリゴ(dT)−開始 λgt10ライブラリー中におけるproHRGβ2とproHRGβ3とを同定した。4つのHRGのそれぞれをコード化する二つのcDNAクローンを配列決定した(エフ・サンガー(F.Sanger),エス・ミルケン(S.Milken),エイ・アール・カルソン(A.R.Coulson),Proc.Natl.Acad.Sci.U.S.A.74,5463 1977))。別のcDNA指定クローン84は、アミノ酸数420を通じてproHRGβ2と同一のアミノ酸配列を有している。位置421の終止コドンの後には、異なった3’−非翻訳配列が続く。
【0184】
例9
別のスプライシング変異体の分離
例6の方法によって、スプライシング変異の結果として4つの密接に関連した配列(ヘレグリンα,β1,β2,β3)が作り出された。ペレス(Peles)他(Cell 69,205(1992))及びウェン(Wenn)他(Cell 69,559(1992))は、p185erbBに結合するタンパク質に関する例1〜4及び6に記載の方法に類似の精製方法とクローン化方法を使用して別のスプライシング変異体を分離した(ラットから)。cDNAクローンは次にようにして得られた(形質転換ラット線維芽細胞系からのp185erbB2結合タンパク質の精製と配列決定による)。
【0185】
p185erbB結合タンパク質は、以下のようにして調整培地から精製された。500のローラボトル(全部で120リットル)の3回の収穫からのプールされた調整培地を、0.2μ フィルターによる濾過によって浄化し、20kd分子径カットオフの膜によるPelicon限外濾過システムによって31倍に濃縮した。すべての精製工程は、Pharmacia fastタンパク質液体クロマトログラフィーシステムを使用して行われた。前記濃縮物を、直接、ヘパリン−セファローズ(150ml,燐酸緩衝食塩水(PBS)によって予め平衡化しておいたもの)に負荷した。カラムを、280nmの波長において全く吸収が検出されなくなるまで、0.2M NaClを含有のPBSで洗浄した。結合タンパク質を、次に、NaCl(250ml)の連続グラジエント(0.2Mから1.0M)で溶出し、5mlのフラクションを採取した。サンプル(前記採取フラクションの0.01ml)を使用して、キナーゼ刺激活性の定量分析を行った。三つのカラム・ラン(全容量=360ml)からの活性フラクションを、プールし、YM10限外濾過膜(アミコン(Amicon),マサチューセッツ州,ダンヴァース)を使用して25mlに濃縮し、1.7Mの濃度となるように硫酸アンモニウムを添加した。遠心分離処理(10,000xg,15分間)による浄化後、プールされた物質をフェニール・スペローズカラム(HR10/10,Pharmacia)上に負荷した。前記カラムを、0.1MのNa2PO4(pH7.4)中で(NH4)2SO4の45mlグラジエント(1.7Mから無塩状態まで)で展開し、2mlのフラクションを採取してそのキナーゼ刺激を分析(一サンプルにつき0.002ml)した(例6に記載のように)。活性の主なピークをプールし、50mMの燐酸ナトリウムバッファ(pH7.3)に対して透析した。一つのモノ−S陽イオン交換カラム(HR5/5,Pharmacia)を、50mMの燐酸ナトリウムで予め平衡させた。活性物質(タンパク質の0.884mg;35ml)の負荷後、前記カラムを、開始バッファで洗浄し、次に、NaClのグラジエントで1ml/分の速度で展開した。キナーゼ刺激活性は、0.45〜0.55M塩で回復され、それぞれ2mlずつの4つのフラクションに渡っていた。
【0186】
これらをプールし、Cu+2キレート化カラムに直接負荷した(1.6ml,HR2/5キレート化スペローズ,Pharmacia)。大半のタンパク質は樹脂に吸着されたが、これらは徐々に塩化アンモニウムの30mlのリニアグラジエント(0〜1M)によって溶出した。前記活性物質は、0.05ないし0.2M NH4Clの範囲の一つのタンパク質ピークで溶出した。様々な精製工程から得られたサンプルを、ゲル電気泳動によって分離し、ICN(カリフォルニア州,コスタ・メサ)のキットを使用した銀染色後、それらのタンパク質内容を、Bio−Red(カリフォルニア州,リッチモンド)のキットを使用したコマーシブルー染色結合分析によって調べた。
【0187】
前記p44タンパク質(10μg)を、0.1Mアンモニウム重炭酸塩バッファ(pH7.8)200μl中で再構築した。L−1−トシル−アミド2−フェニールエチルクロロメチルケトン処理トリプシン(Serva)による消化を、1:10の酵素対基質比で、37℃で18時間かけて行った。その結果得られたペプチド混合物を、Vydac C4マイクロカラム(2.1mm i.d.x15cm,300Å)と、ダイオードアレイ検出器とワークステーションとを備えたHP1090液体クロマトグラフィシステムとを使用し、215nmでモニタしつつ、逆相HPLCで分離した。前記カラムを、0.1%トリフルオロ酢酸(移動相A)で平衡させ、0%から55%の移動相B(0.1%のトリフルオロ酢酸中に90%のアセトニトリル)のリニアグラジエントで70分間にわたり溶出を行った。その流量は0.2ml/分で、カラム温度は、25℃に制御された。前記HPLCシステムから手作業によって採取されたペプチドのピークの1/3部分は、Edman分解によるN−末端配列分析によって特徴付けられた。27.7分後(T27.7)に溶出されたフラクションは、混合アミノ酸配列を有し、還元後、つぎのようにして更に、再クロマトグラフされた。前記ペプチドフラクションの70%の部分を真空中で乾燥し、100μlの0.2M重炭酸塩アンモニウムバッファ(pH7.8)中で再構築した。前記溶液にDTT(最終濃度2mM)を添加し、これを次に37℃で30分間培養した。次に還元したペプチド混合物を、Vydacカラム(2.1mm i.d×15cm)を使用して逆相HPLCによって分離した。溶出条件と流量は、前述したものと同じであった。前記ペプチドのアミノ酸配列の分析を、オンライン・フェニルチオヒダントイン(PTH)アミノ酸分析器を備えたモデル477タンパク質シーケンサ(アプライド・バイオシステムズ・インコーポレイテッド(Applied Biosystems,Inc.),カリフォルニア州,フォスター・シティ)と、モデル900データ分析システム(フンカピラー(Hunkapiller)他(1986)In Methods of Protein Microcharacterization,J.e.Shively,ed.(ニュージャージー州,クリフトン:Humana Press p.223−247)とによって行った。前記タンパク質を、ポリブレンとNaClとでプレサイクルしたトリフルオロ酢酸−処理グラスファイバーディスクに負荷した。PTH−アミノ酸分析を、デュアルシュリンジポンプと逆相(C−18)小口径カラムを使用したマイクロ液体クロマトグラフィシステム(モデル120)で行った(アプライド・バイオシステムズ(Applied Biosystems),2.1mm x250mm)。
【0188】
RNAを、標準手順(マニアティス(Maniatis)他,Molecular Cloning:A Laboratory Manual(Cold Spring Harbor,New York(1982))によってラット1−EJ細胞から分離し、ポリ(A)+をrRNA分離キット(クロンテック・ラボ・インコーポレイテッド,カリフォルニア州,パロ・アルト)を使用して選別した。cDNAをSuperscriptキット(ビー・アール・エル・ライフ・テクノロジーズ・インコーポレイテッド(BRL Life Technologies,Inc.),メリーランド州,ベテスダ)で合成した。カラム−細分化−二重ストランドcDNAをSal1及びNot1−消化pJT−2プラスミドベクター、pCD−Xベクターの派生物に結合し(オカヤマ(Okayama)およびベルク(Berg),Mol.Cell Biol.3:280(1983))、電気泳動によってDH10B E.coli細胞に形質転換した(ダウアー(Dower)他、Nucl.Acids Res.16:6127(1988))。約5x105の一次形質転換物が、N−末端のNDF(残基5〜24)とT40.4トリプシンペプチド(残基7−12)のタンパク質配列から由来の二つのオリゴヌクレオチドプローブによってスクリーニングされた。これらのそれぞれの配列は以下の通りである(Nはすべての4nt示す):
【0189】
前記合成オリゴヌクレオチドを、T4ヌクレオチドキナーゼにより、[γ−32P]ATPによって末端標識化し、これらを使用してニトロセルロースフィルターの複製セットをスクリーニングした。そのハイブリダイゼーション溶液は、6xSSC,50mMの燐酸ナトリウム(pH6.8)と、0.1%のピロ燐酸ナトリウム、2×デンハルト溶液、50μg/ml鮭精子DNAと、20%のフォルムアミドを含有(プローブ1について)又はフォルムアミド0%(プローブ2について)を含んでいた。フィルターを、50℃で0.5×SSC,0.2%ASA,2mM EDTAによって(プローブ1について)、又は37℃で2xSSC,0.2%SDS,2mM EDTAによって(プローブ2について)、洗浄した。これらフィルターのオートラジオグラフィは、両方のプローブとハイブリダイズした10のクローンを示した。これらのクローンを、前述のような再プレート化とプローブハイブリダイゼーションによって精製した。アプライド・バイオシステムズ(Applied Biosystems)373A自動DNAシーケンサとApplied Biosystems Taq DyeDoxy(登録商標)ターミネータサイクル配列決定キットとを、該製造業者の指示に従って使用して、前記cDNAクローンの配列決定をした。いくつかの場合においては、配列は、[35S]dATP(Amersham)及びU.S.BiochemicasのSequenase(登録商標)キットを使用することによって得られた。cDNAクローン44の両方のストランドは、合成オリゴヌクレオチドをプライマーとして使用することによって配列決定された。ほぼ5’350ntの配列が、7つの独立したcDNAクローンにおいて決定された。その結果得られたクローンは、図30示すパターンを示した(NDF)。
【0190】
例10
他の可能なスプライシング変異体の検出戦略
ウシのcDNAクローンとPCR生成物の推定アミノ酸配列、及び公表されているヒト(図31)とラット配列との整合性は、高いレベルの類似性を示して、これはこれらの配列が三つの種の中の相同遺伝子由来のものであることを示している。cDNA/PCR生成物レベルにおいて検出可能なメッセンジャRNA転写体の数が変化するのは、恐らく、広範囲な組織−特異スプライシングに依るものと思われる。得られた、そして図30に示すパターンは、その他のスプライシング変異体が存在することを示唆している。可能なスプライシング変異体のリストが図37に示されている。これらの変異体の多くは、様々な組織から由来のcDNAライブラリーのコード化セグメント特異なプローブ化と、特定のコード化セグメントに特異的なプライマー対を使用したPCR実験とによって得ることが出来る。あるいはこれに代えて、これら変異体は、当業者の公知の切断及びスプライス技術により、特異なcDNAクローン、PCR生成物又はゲノムDNA領域から組み立てることも可能である。例えば、共通のコード化セグメント(例えば、A)中の稀な制限酵素切断サイトを使用して、GGF2BPP5のFBAアミノ末端を、GGF2BPP1、GGFBPP2、GGFPP3、又はGGFPP4のカルボキシ末端配列に接続することができる。コード化セグメントE及び/又はGの存在又は不在が、目的とし、又既述の用途のために有利である場合には、これらのコード化セグメントを、発現構築体に含ませることができる。これらの変異体配列は、組替えシステム中に発現させることができ、その組替え生成物を分析して、そのシュワン細胞細胞分裂誘発活性のレベルと、そのp185erbB2レセプターに結合しこれを活性化する能力とを測定することができる。
【0191】
例11
GGFの機能要素の同定
GGF配列のファミリの推定構造は、その最長の形状(GGF2BPP4によって表される)が、細胞外部分が上皮細胞成長因子に類似するドメインを含む場合に、経膜タンパク質をコード化することを示しているCarpenter and Wahl in Peptide Growth Factors and Their Receptors I pp.69−133,Springer−Verlag,NY 1991参照)。コード化セグメントC及びC/D又はC/D’ペプチド配列におけるシスチン残基の位置は、上皮細胞成長因子(EGF)ペプチド配列に於ける相同残基に対して保存されている(図35、配列識別番号151−153参照)。これは、前記細胞外ドメインが、レセプター認識及び生物学的活性サイトとして機能することを示唆している。変異体形状の内のいくつかは、H,K及びLコード化セグメントを欠如しており、従って、分泌、拡散可能生物学的活性タンパク質として発現可能である。EGF状ドメイン(EGFL)を含むポリペプチドをコード化するGGF DNA配列は、グリア細胞細胞分裂誘発活性を刺激するための十分な生物学的活性を有している可能性がある。
【0192】
このタンパク質の膜結合バージョンは、胚形成中又は、神経再生中(ニューロンの表面が増殖シュワン細胞の表面に密接に関連している場合)にニューロンの表面に発現された場合、シュワン細胞の増殖を誘発する可能性がある。
【0193】
分泌(非膜結合)GGFは、シュワン細胞とその分泌位置からいくらか離れた位置において作用することが可能な分類上拡散可能な因子として作用する可能性がある。他の形態も、細胞の損傷や細胞の破壊を通じた源から細胞内から放出される可能性がある。分泌GGFの一例は、GGF2HBS5によってコード化されるタンパク質であり(例6参照)、これは細胞の外部に向けられることが判った唯一のGGFである(例7)。分泌は、恐らく、GGF2HBS5によってコード化される組替えGGF−II中に含まれるN−末端ドメインであって、領域Eにおいてのみ見られるN−末端疎水性配列によって仲介される。
【0194】
他のGGFは、分泌されるものではないように思える(例6参照)。これらのGGFは、組織の損傷の結果として放出される損傷反応形態であるのかもしれない。
【0195】
GGF−II(GGF2HBS5によってコード化される)の予想タンパク質構造の他の領域と、領域B及びAを含む他のタンパク質とは、ヒト基底膜ヘパラン硫酸プロテオグリカン核タンパク質(ref.)に対する類似性を示す。これらのGGFにおけるC2免疫グロブリンフォールドの第2システインの隣に位置するペプチドADSGEYは、基底薄膜タンパク質に見られる22のC−2反復の内の9つに発生する。この証拠は、これらのタンパク質が、ニューロンやグリア等に関連するマトリックスタンパク質に関連していることを強く示唆するものであり、その目標サイトとしてのグリア成長因子の隔離の方法を示唆している可能性がある。
【0196】
例12
組替え細胞からのGGFの精製
その生物学的活性を分析するべく完全長GFFあるいはGGFの部分を得るために、タンパク質を、クローン化DNAを使用して過剰生産することができる。いくつかの方法が使用可能である。前述の配列を有する組替えE.coli細胞を構築することができる。pNH8aやpHH16a(ストラトジェン・インコーポレイテッド(Stratagne,Inc.))等の発現システムをその製造業者の指示に従ってこの目的のために使用することができる。あるいはこの代わりに、これらの配列を、ほ乳類発現ベクターに挿入して、過剰生産細胞系を構築することができる。例えば、この目的のために、GGFをコード化するDNA、クローンGGF2BPP5を、COS細胞と、チャイニーズハムスタの卵巣細胞との両方に発現させた(例7参照)(J.Biol.Chem.263,3521〜3527(1981))。このGGGF DNAを有するベクターを、公知の手法を使用してホスト細胞に移入することができる。
【0197】
一過的発現を調べるか、もしくは、G418耐性クローンを、メトトレキセートの存在下おいて成育させることにより、dhfr遺伝子(pMSXNDベクター上に含まれる)を増幅し、その過程において、隣接のGGFタンパク質コード化配列を共増幅する細胞を選別することができる。全く血清やタンパク質を含まない培地(ハミルトン(Hamilton)およびハム(Ham),In Vitro 13,537〜547(1977))中に、CHO細胞維持することができるので、所望のタンパク質は、その培地から精製することが可能である。例9において作られた抗血清を使用したウェスタン分析を使用して、前記過剰生産細胞の調整培地中における所望のタンパク質の存在を検出することができる。
【0198】
所望のタンパク質(rGGF−II)を、次のようにして、COS細胞を一過的に発現させることによって調整した培地から精製した。rGGF−IIを、調整培地から収穫し、陽イオン交換クロマトグラフィ(POROS−HS)を使用して部分精製した。前記カラムを、33.3mM MES pH6.0で平衡化した。調整培地を、10ml/分の流量で負荷した。シュワン細胞増殖活性と免疫反応性(ポリクローナル抗血清を上述のようにGGFIIペプチドに対して使用した)とを有するピークを、50mM Tris,1M NaCl pH8.0で溶出した(それぞれ、図50A及び50B)。
【0199】
rGGF−IIも、安定したチャイニーズハムスタの卵巣細胞系を使用して発現される。収穫された調整培地からのrGGF−IIを、陽イオン交換クロマトグラフィ(POROS−HS)を使用して部分精製した。前記カラムを、PBS pH7.4によって平衡化した。調整培地を、10ml/分で負荷した。シュワン細胞増幅活性と免疫反応性(GGF−IIポリクローナル抗血清を使用)とを有するピークを、50mM Hepes,500mM NaCl pH8.0で抽出した。増殖性と免疫反応性の両方を備えた別のピークが、50mM Hepes,1M NaCl pH8.0で観察された。
【0200】
rGGF−IIは、高分解能工程として疎水性反応クロマトグラフィ(陽イオン交換/リザーブフェーズクロマトグラフィ(必要な場合には第2高分解能工程として)、陰イオン交換クロマトログラフィのようなウィルス不活性化工程及びDNA除去工程、を使用することによって更に精製することができる。
【0201】
使用した手法の詳細な記述は以下の通りである。
【0202】
前記陽イオン交換カラムから抽出した組替えGGF−IIピークのシュワン細胞増幅活性は、以下のようにして測定した。培養シュワン細胞の細胞分裂誘発反応を、50mM Tris 1M NaCl pH8.0によって抽出したピークを使用して5Mフォルスコリンの存在下において測定した。前記ピークは、20 1,10 1(1:10)10 1及び(1:100)10 1で添加した。125I−ウリジンの組み込みを測定し、18〜24時間の露光後に(CPM)として表現した。
【0203】
GGF−IIのペプチドに対して発育させたポリクローナル抗体を使用した免疫ブロットを、次のようにして行った。10μlの異なったフラクションを、4−12%のグラジエントゲルでランさせた。これらのゲルを、ニトロセルロース紙に移し、このニトロセルロースブロットを5%のBSAでブロックし、GGF−II−特異抗体(1:250希釈)でプローブした。125Iタンパク質A(1:500希釈、特異的活性=9.0/Ci/g)を、二次抗体として使用した。前記免疫ブロットを、コダック社X線フィルムに6時間露光した。1M NaClで溶出したピークフラクションは、65〜90Kdにおいて、GGF−IIと高分子量グリコフォームの予想サイズ範囲である,広い免疫反応性バンドを示した。
【0204】
陽イオン交換カラムでのGGF−II精製は以下のように行った。rGGFIIを発現するCHO細胞調整培地を、10ml/分で前記陽イオン交換カラムに負荷した。このカラムを、PBS pH7.4で平衡化した。溶出は、それぞれ、50mM Hepes 500mM NaCl pH8.0と50mM Hepes 1M NaCl pH8.0とで行った。すべてのフラクションを、ここに記載のシュワン細胞増殖分析(CPM)を使用して分析した。タンパク質濃度(mg/ml)は、BSAを標準として使用したBradford分析によって測定した。
【0205】
10μlの各フラクションを使用したウェスタンブロットを行った。図51A及び51Bに示すように、免疫反応性とシュワン細胞活性とは相互移動する。
【0206】
前述のシュワン細胞細胞分裂誘発分析を使用して、完全長クローン又はその全ての物学的活性部分からの発現生成物の分析を行った。完全長クローンGGF2BPP5が、COS細胞に一過的に発現された。移入されたCOS細胞の細胞内抽出物は、例1に記載したシュワン細胞増殖分析により分析した時に生物学的活性を示す。更に、GGF2HBS5を発現する完全長に近いものが、CHOと昆虫(例7)細胞とに一過的に発現された。この場合、細胞抽出物と調整培地との両方が、例1に記載したシュワン細胞増殖分析において生物学的活性を示す。当業者によって、GGF(ヘレグリンを含む)からのスプライシング変異体相補性DNAのファミリのすべてのメンバを、このようにして発現させ、シュワン細胞増殖分析において分析することができる。
【0207】
あるいは、組替え物質を、COS−7細胞においてスプライシング変異体Neu分化因子(NDF)を発現させたウェン(Wen)他(Cell 69,559(1992))に従って、他の変異体から分離することができる。pJT−2真核生物プラスミドベクターに挿入したcDNAクローンは、SV40初期プロモーターのコントロール下にあり、SV40終結及びポリアデニール化信号に3’で挟まれている。COS−7細胞は、以下のようにして、電気ポレーションによって、pJT−2プラスミドDNAで移入された。即ち、6x106の細胞を(0.8mlのDMEMと10%のFEBSにおいて)、0.4cmのキュベットに移し、10μLのTE溶液中(10mM Tris−HCl(pH8.0),1mM EDTA)にて20μgのプラスミドDNAと混合した。エレクトロポレーションは、パルスコントローラユニットを200オームに設定したBio−Rad Pulser装置を使用して、室温で、1600V、24μFで行った。次に、細胞を、20mlのDMEM,10%FBS中に希釈し、T75フラスコ(Falcon)中に移した。37℃における14時間の培養後、培地を、DMEM,1%FBSと置換し、培養を更に48時間継続した。細胞から収穫した組替えタンパク質を含む調整培地は、このタンパク質のレセプターを発現する細胞系において生物学的活性を示した。この細胞系(ヒト乳ガン細胞ラインAU565)を、組替え物質で処理した。処理された細胞は、erbB2レセプターの活性化に特徴的な形態変化を示した。このタイプの調整培地も、シュワン細胞増殖分析においてテストすることが出来る。
【0208】
例13
p185レセプターに結合する他のタンパク質の精製と分析
I.gp30とp70との精製
ここに参考文献として添付するルプ(Lupu)他(Science 249,1552(1992))及びリップマン(Lippman)およびルプ(Lupu)(特許出願番号PCT/US91/03443(1990))は、以下のようにして、ヒト乳ガン細胞ラインMDA−MB−231の調整培地からタンパク質を精製した。
【0209】
調製培地採取を、周知の方法を使用して行った。培地を、Amicon限外濾過フィルターセル(YM5膜)(Amion,Danvers,MA)中にて100倍に濃縮した。浄化し濃縮した後、培地を、−20℃で保存し、一方、連続採取を以後数日の間に行った。濃縮培地を、4℃で2日間に渡り、100容量の0.1Mの酢酸に対して、Spectra/por3チュービング(スペクトラム・メディカル・インダストリーズ(Spectrum Medical Industries),カリフォルニア州,ロス・アンジェルス)を使用して透析した。透析中に沈澱した物質を、4℃で30分間、4000rpmでの遠心分離処理によって除去し、プロテアーゼインヒビターを添加した。次に、浄化したサンプルを凍結乾燥した。
【0210】
凍結乾燥した調整培地を、全タンパク質が約25mg/mlの最終濃度となるように、1Mの酢酸中に、溶解させた。不溶性物質は、10,000rpmで15分間の遠心分離処理によって除去した。次に、このサンプルをセファーデックスG−100カラム(XK16,Pharmacia,Piscataway,NJ)に負荷し、平衡化し、更に、これに、4℃で1Mの酢酸を30ml/時間の流量で上方に流すことによる溶出を行った。4mlの100倍濃縮培地から、100ngのタンパク質が処理された。3mlの溶出物を含むフラクションを、凍結乾燥させ、分析用に300μlのPBS中で再懸濁させ、更なる精製のための原料として利用した。
【0211】
セファデックスG−100精製物を、逆相高圧液体クロマトグラフィ(HPLC)でランした。第1の工程には、急勾配のアセトニトリルグラジエントを使用した。急勾配アセトニトリルグラジエントと他のすべてのHPCL工程とは、C3−逆相カラムを、水中(HPLC−グレード)0で0.05%のTFA(トリフルオロ酢酸)によって平衡化した後に、室温で行った。これらのサンプルを負荷し、フラクションは、1ml/分の流量で30分間に渡って、リニアグラジエント(0.05%のTFA中で0〜45%のアセトニトリル)で溶出した。280nmで吸収が観察された。1mlのフラクションを採取し、EGFレセプター競合活性の分析の前に凍結乾燥させた。
【0212】
第2のHPLC工程には、緩勾配アセトニトリルグラジエントを使用した。前のHPLC工程からの活性フラクションのプールを、同じカラムに関して再びクロマトグラフィ分析した。0.05%のTFA中で5分間の0−18%アセトニトリルグラジエント後に、0.05%のTFA中で30分間のリニアな18〜45%アセトニトリルグラジエントにて、溶出を行った。流量は1.0ml/分で、1mlのフラクションが採取された。ヒトTGF瘁|状因子が、30〜32%のアセトニトリル濃度で、RRAによる検出可能な一つのピークとして溶出した。
【0213】
ルプ(Lupu)他(Proc.Natl.Acad.Sci.89,2287(1992))は、p185erbB2レセプターに結合する別のタンパク質を精製した。この特定のタンパク質、p75、は、10%の胎児ウシ血清(GIBCO)を補足した改良イーグル培地(IMEM:GIBCO)中で繁殖させたSKBr−3(ヒト乳ガン細胞系)の成長に使用される調整培地から精製されたものである。タンパク質p75は、p185erbB2アフィニティーカラムを使用した濃縮(100倍)調整培地から精製された。p185erbB2の94キロダルトン細胞外ドメイン(p75に結合)を、組替え発現によって生成し、ポリアクリルアミドヒドラジド−セファローズアフィニテイークロマトグラフィマトリックスに結合させた。結合後、前記マトリックスを、氷冷した1.0M HCl中で十分に洗浄し、そのビーズを0.5M NaNO2で活性化させた。温度は20分間、0℃に維持し、その後に、濾過と、氷冷した0.1M HClでの洗浄を行った。500mlの濃縮調整培地を、重力によって前記ビーズに流した。カラムを洗浄し、1.0Mのクエン酸で、4.0から2.0のpH値で(erbB2とp75を分離させるため)ステップ的に溶出させた。すべてのフラクションを、Pharmacia PD10カラム上で脱塩させた。精製によって、3.0−3.5溶出pHで75kDaの均−なポリペプチドが得られた(銀染色によるSDS/PAGEでの分析によって確認)。
【0214】
II.gP30のP185 erbB2 B2への結合
精製gp30タンパク質を、それがp185erbB2に結合したか否かを調べる分析においてテストした。p185erbB2に対するモノクローナル抗体を備えた競合アッセイ。gp30タンパク質は、SK−BR−3及びMDA−MB−453細胞(p185erbB2レセプターを発現するヒト乳ガン細胞系)中でP185erbB2に結合する抗体を置換した。gp30のシュワン細胞増殖活性は、更に、シュワン細胞培養物を、例1〜3に記載の分析手法を使用して、精製pg30によって処理することによっても示すことができる。
【0215】
III.p75のp185 erbB2 への結合
SKBr−3調整培地から得た75kDaのポリペプチド(p75)が実際に、SKBr−3細胞中のerbB2ガン・タンパク質のためのリガンドであるのか否かを分析するために、gp30のための前述の競合アッセイを使用した。p75が結合活性を示すことが判ったが、これに対して、他のクロマトグラフィフラクションからの物質はそのような活性を示さなかった(データは図示せず)。通過物質は、いくらかの結合活性を示した。これは、抜け出たerbB2 ECDの存在によるものであろう。
【0216】
IV.他のp185 erbB2 リガンド
ペレス(Peles)他(Cell 69,205(1992))は、更に、ラット細胞(NDF,その方法については例8参照)からのp185erbB2刺激リガンドをも精製した。ホルムズ(Holmes)他(Science 256,1205(1992))は、p185erbB2に結合し、これを刺激する、ヒト細胞からのヘレグリンαを精製した(例6参照)。ここに参考として添付する、タラコフスキー(Tarakovsky)他Oncogene 6:218(1991)は、活性化マクロファージから分離された25kDのポリペプチドの、p185erbB2相同体である、Neuレセプターへの結合を示した。
【0217】
VI.NDFの分離
ヤーデン(Yarden)およびペレス(Peles)(Biochemistry 30,3543(1991))は、p185erbB2レセプターを刺激するであろう35キロダルトンのグリコタンパク質を同定した。このタンパク質は、次の手法に従って調整された培地中において同定された。ラットI−EJ細胞を、175−cm2フラスコ(Falcon)中で集合成長させた。単層を、PBSで洗浄し、無血清培地中にて10〜16時間放置した。前記培地を廃棄し、新たな無血清培地によって置換し、3日間の培養後に採取した。調整培地を、低速遠心分離によって浄化し、YM2膜(分子量2000でカットオフ)を備えたAmicon限外濾過セル中で100倍に濃縮した。調整培地におけるneu刺激活性の生化学的分析は、前記リガンドが、熱に対しては安定しているが還元に対しては敏感な35−kDグリコタンパク質であることを示している。該因子は、高い塩濃度か、酸性アルコールのいずれかによって沈澱させることができる。選択的沈澱、ヘパリン−アガロースクロマトグラフィ、及び希釈酸中でのゲル濾過によって前記分子を部分精製することにより活性リガンドが得られ、これはがん原遺伝子のレセプターを刺激することが出来るが、構造的に活性ながん遺伝子neuタンパク質に対しては無効果であった。しかしながら、この精製されたフラクションは、EGFのための関連レセプターをも刺激する能力を保持しており、これはこれらの二つのレセプターが、双方向メカニズムを介して機能的に結合していることを示唆するものである。あるいは、前記推定リガンドは、同時に二つのレセプターと反応する。該因子の、提示された生化学的特徴を利用して、これらの可能性が探求される完全に精製された因子を達成することができる。
【0218】
他の出版物において、デイヴィス(Davis)他(Biochem,Biophys.Res.Commun.179,1536(1991),Proc.Natl.Acad.Sci.88,8582(1991)およびグリーン(Green)他,PCT特許出願PCT/US91/02331(1990))は、ヒトT−細胞(ATL−2)系の調整培地からのタンパク質の精製について記載している。
【0219】
ATL−2細胞系は、IL−2独立HTLV−1(+)T細胞系である。無マイコプラズムATL−2細胞を、5%のCO2を含む加湿雰囲気中において37℃で、培養培地として10%のFCBを含有するRPMI 1640(10%FCS−RPMI 1640)培地中に保持した。
【0220】
タンパク質物質の精製のために、ATL−2細胞を、1xPBSで2回洗浄し、72時間、無血清RPMI 1640培地/2mM L−グルタミン中で3x105mlで培養し、その後、前記細胞をペレット化した。このように生成した培養上澄を、「調整培地」(C.M.)と称する。
【0221】
C.M.を1000dカットオフのYM−2Diaflo膜(アミコン(Amicon),マサチューセッツ州,ボストン)を使用して、1リットルから10mlへと濃縮した。いくつかの分析用に、1000MW以上の成分を含有する濃縮C.M.を、RPMI培地によって、もとの容量に再希釈した。ポリアクリルアミドグラジエントゲル(インテグレイテッド・セパレーション・システムズ(Integrated Separation Systems),メリーランド州,ハイド・パーク又はフォアキャスト・システム・バイ・アマーシャム(Phorecast System by Amerscham),イリノイ州,アーリントン・ハイツ)を使用してゲル電気泳動にかけ、その後、前記1リットルの調合物からのこの2つのカラム精製物資の幾らかを銀染色したところ、少なくとも4ないし5のバンドが現れ、これらの内、10kDと20kDとのバンドがこの物質に特有のものであった。通過した、1000NW以下のC.M.含有成分は希釈せずに使用した。
【0222】
濃縮調整培地を、.45μ ユニフロフィルター(Schleicher and Schuell,Keene,NH)でフィルター殺菌し、次に、予め10mM Tris−Cl,pH8.1で平衡化させておいたDEAE−SW陰イオン交換カラム(ウォーターズ・インコーポレイテッド(Waters,Inc.),マサチューセッツ州,ミルフォード)への適用によって更に精製した。一ランのHPLC当り1リットルの元のATL−2調整培地を表す濃縮C.M.タンパク質を、前記カラムに吸着させ、次に、4ml/分の流量で、0mMから40mMのNaClのリニアグラジエントで溶出した。フラクションを、10%の適当なDEAEフラクション(1カラム精製物)又は1%の適当なC18フラクション(2カラム精製物)に対する生体外免疫複合体キナーゼ分析を使用して、分析した。この生体外免疫複合体キナーゼ分析を使用して投与量−依存的にp185c−neuのチロシンキナーゼ活性を増加させた活性物質は、220ないし240mMのNaClの近辺での4ないし5のフラクション(36−40)に渡って一つの主要ピークとして溶出した。HPLC−DEAE精製後、前記活性フラクション中のタンパク質を、濃縮、プールして、更に濃縮し、その後、C18(百万マトリクス)逆相クロマトグラフィ分析した(ウォーターズ・インコーポレイテッド(Waters,Inc.),マサチューセッツ州,ミルフォード)(C18+1工程または2カラム精製物と称する)。溶出は、0.1%のTFAに対する2−プロパノールのリニアグラジエントで行った。すべてのフラクションを、RPMI1640培地に対して透析して2−プロパノールを除去し、下記のように、前記生体外免疫複合体キナーゼ分析と、適当なフラクションの1%濃縮物を使用して分析した。p185c−neuのチロシンキナーゼ活性を増加させる活性物質は、二つのピークで抽出された。一つはフラクション11〜13において溶出し、第2番目の僅かに低い活性の活性物質ピークは、フラクション20〜23において溶出した。これらの二つのピークは、それぞれ、約5ないし7%のイソプロパノールと約11ないし14%のイソプロパノールとに対応している。C18#1発生フラクション11〜13を、特徴付け研究に使用した。第2クロマトグラフィ工程から得られた活性フラクションをプールし、タンパク質物質サンプルに指定した。
【0223】
20リットルの調合も同じ精製戦略を使用した。前記DEAE活性フラクション35〜41をプールし、前述のようにしてc18クロマトログラフィ分析にかけた。C18#1フラクション11〜13と21〜24との両方が、投与量−依存活性を有していた。フラクション11〜13のプールを、更にC18クロマトグラフィ分析工程にかけた(C18#2又は3カラム精製物と称する)。ここでも再び、フラクション11〜13と21〜24とが活性を有していた。例8で記載の生体外免疫複合体キナーゼ分析において測定されたフラクション23の投与量反応は、フラクション23を容量で0.005%、フラクション23を容量で0.05%添加することによって得られる。これは達成された最も高い純度である。
【0224】
分子量の範囲を、ゲル濾過クロマトグラフィ分析と限外濾過膜分析とに基づいて測定した。ほぼ同じ量のチロシンキナーゼ活性を、保持し、10,000分子量カットオフフィルターにより通過した。ほとんどすべての活性物質が、30,000分子量カットオフフィルターを通過した。活性物質クロマトグラフフラクションの分子量範囲を、投与量−依存neu−活性化活性物質を含むフラクションを、同じ実行条件を使用して発生された一組のタンパク質分子量標準(シグマ・ケミカル・カンパニー(Sigma Chemical Co.,ミズーリ州,セント・ルイス)の溶出プロフィールと比較することによって決定した。活性物質の低分子量の領域が、7,000と14,000ダルトンの間に見つかった。活性物質の第2の範囲は、約14,000から約24,000ダルトンの間であった。
【0225】
ポリアクリルアミドグラジエントゲル(インテグレイテッド・セパレーション・システムズ(Integrated Separation Systems,メリーランド州,ハイド・パーク又はフォアキャスト・システム・バイ・アマーシャム(Phorecast System by Amerscham),イリノイ州,アーリントン・ハイツ)を使用した電気泳動後、3−カラム精製物質(c18#2)の銀染色を、市販の銀染色キット(BioRad,Rockville Centre,NY)を使用して行った。20リットルの調合物のc18#2精製から得られたフラクション21,22,23及び24を、マーカーとともにランさせた。フラクション22と23とが、185erbB2(neu)キナーゼ分析(下記参照)において最も強力な投与量反応を示した。選択された分子量フラクションがp185erbB2と作用するという事実が、免疫複合体キナーゼ分析によって示された。
【0226】
ここに参考文献として添付するフアング(Huang)他(1992,J.Biol.Chem.257:11508〜11512)は、ウシの腎臓から別のneu/erb B2リガンド成長因子を分離した。25kDのポリペプチド因子が、カラム分別の手法によって分離され、次いで、DEAE/セルロース(DE52)、Sulfadex(硫酸エステル化されたセファーデックスG−50)、ヘパリン−セファロース4B、そしてSuperdex75(高速タンパク質液体クロマトグラフィ)で連続カラムクロマトグラフィー分析された。前記因子、NEF−GF、は、neu/erbB2遺伝子生成物のチロシン−特異性自己燐酸化を刺激する。
【0227】
VII.p185 erbB2 B2に結合するリガンドに対する免疫複合体分析NDF
この分析は、ATL−2調整培地(C.M.)又はタンパク質物質を変量させながらPN−NR6細胞溶解物の予備培養によって推進した、免疫沈降したp185の自己燐酸化活性物質における相違を示すものであって、これをここでneu−活性化活性物質と称する。
【0228】
免疫複合体キナーゼ分析において使用した細胞系は、ここにその全部を参考文献として添付する、コカイ(Kokai)他,Cell 55,287〜292(July 28,1989)と、これもその全部を参考文献として添付する、マーク・アイ・グリーン(Mark I.Green)の名のもとに「抗−レセプター抗体によってがん細胞を治療する方法」という名称で1989年7月27に出願された米国出願第386,820号とに記載の方法によって、入手、調合、培養したものである。
【0229】
細胞ラインは、すべて、5%のCO2を有する加湿雰囲気中において、培養培地として5%のFCSを含有するDMEM培地(5%FCS−DMEM)中に維持した。
【0230】
150mmディシュ中の細胞の濃縮培養物を、冷たいPBSで2回洗浄し、10mlの冷凍−解凍バッファ(150mM NaCl,1mM MgCl2,20mM Hepes,pH7.2,10%グリセロール,1mM EDTA,1%アプロチニン)中に掻き入れ、遠心分離(600×6,10分間)した。細胞ペレットを、1mlの溶出(Lysis)バッファ(50mM Hepes,pH7.5,150mM NaCl,3%Brij35,1mM EDTA,1.5mM MgCl2,1%アプロチニン,1mM EGTA,20μlM Na3VO4,10%グリセロール)中で再懸濁させ、4℃で30分間回転させた。
【0231】
すべての化学物質は、特に明記の無い限り、シグマ・ケミカル・カンパニー(Sigma Chemical Co.),ミズーリ州,セント・ルイスからのものであった。非溶解物質を、40,000xgで30分間の遠心分離によって除去した。次に使用するクリアな上澄を、細胞溶出物と指定する。
【0232】
前記細胞溶出物を、50μlの50%(容量/容量)sプロテインA−セファローズ(シグマ・ケミカル・カンパニー(Sigma Chemical Co.),ミズーリ州,セント・ルイス)と、15分間培養し、2分間の遠心分離で、溶出物を予備浄化した。この予備浄化細胞溶出物の50μl部分を、調整培地、タンパク質物質、又は特定の他の因子とともに、溶出バッファと1mlの最終容量で、氷上で15分間培養した。次に、サンプルを、p185neu及びp185c−neuを認識する5μgの7.16.4モノクローナル抗体又は他の適当な抗体と共に、氷上で20分間培養し、その後4℃で回転させながら、50μlの50%(容量/容量)タンパク質A−セファロースで培養した。免疫複合体を、遠心分離によって収集し、500μlの洗浄バッファ(50mM Hepes,pH.7.5、0.1%,Brij35、150mM NaCl、2mM EDTA、1%アプロンチニン、30μm Na3VO4)で4回洗浄し、更に、反応バッファ(20mM Hepes(pH7.4)、3mM MnCl2、及び0.1%Brij35、30μm Na3VO4)で2回洗浄した。ペレットを、50μlの反応バッファ中で再懸濁させ、(ガンマ−32P)−ATP(アマーシャム(Amersham),イリノイ州,アーリントン・ハイツ)を添加して0.2μmの最終濃度を得た。これらのサンプルを、27℃で20分間、又は純粋サンプルとともに4℃で25分間培養した。2mM ATPと2mM EDTAとを含有する3xSDSサンプルバッファを添加することによって反応を終結させ、次に、これらを100℃で5分間培養した。これらのサンプルを、次に、10%のアクリルアミドゲル上でSDS−PAGE分析にかけた。ゲルを染色、乾燥し、強化スクリーンを備えたKodak XARまたはXRPフィルムに露光した。
【0233】
VIII.活性を有するアセチルコリンレセプター(ARIA)の精製
ARIA、即ち、アセチルコリンレセプターの合成を刺激する42kDのタンパク質が、Gerald Fischbach(フォールズ(Falls)他 Cell 72:801−815(1993))の実験室にて分離された。ARIAは、p185erbB2に類似の185Kda筋肉経膜タンパク質のチロシン燐酸化を誘発し、培養胚筋管内のアセチルコリンレセプター合成を刺激する。ARIAをコード化するcDNAクローンの配列分析は、ARIAが、タンパク質のGGF/erbB2リガンドグループのメンバであることを示しており、これは、グリア細胞細胞分裂誘発を刺激や、ここに記載の例えばGGF2の適用に潜在的に有効である可能性を有している。
【0234】
例14
シュワン細胞中のGGFによって仲介されたタンパク質チロシン燐酸化
増殖を誘発するための十分なレベルのグリア成長因子での処理の後、ラットシュワン細胞は、タンパク質チロシンの燐酸化の刺激を示す(図36)。様々な量の部分精製GGFを、例3に概述した手法に従ってラットシュワン細胞の一次培養に適用した。シュワン細胞を、ポリD−リジン被覆した24のウェルプレート内の、1mLのGGF−CMにつき(1ウェル当り0.5mL)DMEM/10%ウシ胎仔血清/5μMフォルスコリン/0.5μg中で発育させた。集合状態において、前記細胞に、ウェル当り0.5mLでDMEM/10%のウシ胎仔血清を供給し、培養器内に一晩放置して鎮静化させた。次の日、前記細胞に、0.2mLのDMEM/10%ウシ胎仔血清を供給し、培養器内で1時間放置した。次に、テストサンプルを、必要に応じて、様々な濃度と長さで、前記培地に直接添加した。次に、細胞を、沸騰している溶解バッファ(燐酸ナトリウム、5mM,pH6.8;SDS,2%,β−メルカプトエタノール、5%;ジチオトレイトール、0.1M;グリセロール、10%;ブロモフェノールブルー,0.4%;バナジウム酸ナトリウム,10mM)中で溶解し、沸騰水浴中で10分間培養し、次に、直接に分析するか、あるいは、−70℃で冷凍した。サンプルは、7.5%SDS−PAGEゲル上でのランによって分析し、トウビン(Towbin)他(1979)Proc.Natl.Acad.Sci.USA76:4350−4354に記載の標準手法を使用してニトロセルロース上にエレクトロブロッティングした。ブロットされたニトロセルロースを、Kamps and Selton(1988)Oncogene 2:305−315に記載されている標準方法を使用して、抗ホスホチロシン抗体でプローブした。プローブされたブロットを、一晩オートラジオグラフィに露光し、標準実験室手法を使用して現像した。濃度測定を、ウルトラスキャンXL強化レーザ濃度計(LKB)を使用して行った。分子量指定は、予備染色高分子量標準(Sigma)に対して行われた。タンパク質燐酸化とシュワン細胞増殖の投与量反応は非常に類似している(図36)。燐酸化バンドの分子量は、p185erbB2の分子量に非常に近い。シュワン細胞を、GGF2HBS5クローンによってCOS細胞翻訳物から調整した培地によって処理した場合にも類似の結果が得られた。これらの結果は、予想されていた、GGFsとの相互作用および185erbB2の活性化とよく相関している。
【0235】
この実験を組替えGGF−IIで繰り返した。GGF−IIクローンで安定的に形質転換したCHO細胞系(GGF2HBS5)からの調整培地は、上述の分析を使用して、タンパク質チロシン燐酸化を刺激する。疑似移入されたCHO細胞系ではこの活性を刺激することは出来なかった(図52)。
【0236】
例15
MDA−MB−231細胞系からのタンパク質因子によるシュワン細胞増殖の分析
シュワン細胞の増殖は、ヒト乳ガン細胞ラインMDA−MB−231由来の調整培地によって媒介される。分析の第1日目、104個の一次ラットシュワン細胞を、96ウェル・ミクロタイター(microtiter)プレート中に、ウェル当り5%のウシ胎仔血漿を補足した100μlのダルベッコ変法イーグル培地内にプレート化した。分析の第2日目、10μlの調整培地(例6に記載のように培養された、ヒト乳ガン細胞ラインMDA−MB−231からのもの)を、前記ミクロタイタープレートの各ウェルに添加した。第6日目、各プレート当りのシュワン細胞の数を、酸ホスファターゼ分析(コノリー(Connolly)他Anal.Biochem.152:136(1986)の手法に従って)によって測定した。前記プレートを、100μlの燐酸緩衝食塩水(PBS)と100μlno反応バッファ(0.1m酢酸ナトリウム,(pH5.5))と0.1%のトリトン(Triton)X−100で洗浄し、各ウェル毎に、10mMの燐酸p−ニトロフェニルを添加した。プレートを37℃で2時間培養し、10μlの1N NaOHを添加することによって反応を終結させた。各サンプルの光濃度を、分光光度計で410nmで読み取った。コントロール細胞系(HS−294T,erbB−2リガンドの非プロデューサ)からの調整培地によって処理したシュワン細胞において細胞数の38%の刺激が観察された。この結果は、MDA−MB−231細胞ライン(p185erbB2結合活性を刺激する)によって分泌されたタンパク質が、シュワン細胞の増殖を刺激することを示すものである。
【0237】
例16
GGFのN−グリコシル化
GGF−II候補クローンGGF2BPP1,2及び3のcDNA配列から予測されるタンパク質配列は、多数のコンセンサスN−グリコシル化モチーフを有している。GGFII02ペプチド配列におけるギャップは、これらのモチーフの一つにおけるアスパラギン残基に一致し、これは、このサイトにおいて恐らく炭水化物が結合していることを示している。
【0238】
GGFのN−グリコシル化を、タンパク質中において炭水化物とアスパラギン残基との間の共有結合を切断する酵素であるN−グリカナーゼとの培養後においてSDS−PAGE上の移動度の変化を観察することによって調べた。
【0239】
GGF−IIのN−グリカナーゼ処理によって、MW40〜42kDaの主要なバンドと、45〜48kDaの副バンドとが得られた。非還元条件下における活性物質溶出実験は、約45〜50kDaにおける一つの活性脱グリコシル化種を示した。
【0240】
GGF−Iの活性物質溶出実験も、N−グリカナーゼによって処理された場合に、MW26〜28kDaの活性種を生ずることにより、電気泳動的移動度の増加を示している。銀染色によって、移動度のシフトがあることが確認されたが、使用されたサンプルのバックグラウンドの染色によってN−脱グリコシル化バンドを指定することが出来なかった。
【0241】
寄託
T7プロモーターのコントロール下におけるプラスミドpブルースクリプト5k中の核酸コード化GGF−II(cDNA,GGF2HBS5)タンパク質(例6)を、アメリカ培養細胞系統保存機関,メリーランド州ロックスビルに寄託し、ATTC番号75298を与えられた。本出願人は、もしもこのプラスミドが、本件が特許となった時にこの特許の権利期間が終結するまでに生存不能となった場合には、これを取り替える義務と、更に、そのような特許が発行されたことをATTCに通知し、その発行時にこの寄託物を公衆に利用可能とする義務があることを認識するものである。それ以前においては、該寄託物は、37CFR 1.14及び35 USC 112の規定に従って特許庁長官に利用可能とされる。
【図面の簡単な説明】
【図1】例1に関し、カルボキシルメチルセルロースクロマトグラフィからの生成物のプロフィール。
【図2】例1に関し、ハイドロキシアパタイトHPLCからの生成物のプロフィール。
【図3】例1に関し、モノ S FPLCからの生成物のプロフィール。
【図4】例1に関し、ゲル濾過処理FPLCの生成物のプロフィール、
【図5】例1に関し、逆相HPLCからの二つの部分的に精製されたポリペプチド生成物のプロフィール。
【図6】例1に関し、逆相HPLCからの二つの部分的に精製されたポリペプチド生成物のプロフィール。
【図7】例1に関し、ウシ胎仔血清又はウシ胎仔血漿のバックグランドを使用した逆相HPLCからのGCF−II及びGCF−IIの投与量−反応曲線。
【図8】例1に関し、ウシ胎仔血清又はウシ胎仔血漿のバックグランドを使用した逆相HPLCからのGCF−II及びGCF−IIの投与量−反応曲線。
【図9】GCF−IとGCF−II,配列認識番号1〜20、22〜29、32〜53及び169から由来のペプチド配列のリスト(例2参照)。
【図10】GCF−IとGCF−II,配列認識番号1〜20、22〜29、32〜53及び169から由来のペプチド配列(後記の例2参照)を示し、特に新規な配列を示し、パネルAは、縮重オリゴヌクレオチドプローブと縮重PCRプライマーとを構成するのに使用するGCF−Iペプチドの配列がリストされ(配列認識番号20,1,22〜29及び17)、パネルAのこれらの配列の内のいくつかは、合成ペプチドの構成にも使用され、パネルBは、縮重プローブと縮重PCRプライマーの構成には短すぎた(6アミノ酸以下)新規なペプチドの配列のリスト(配列認識番号17及び52)。
【図11】GCF−IとGCF−II,配列認識番号1〜20、22〜29、32〜53及び169から由来のペプチド配列(例2参照)を示し、特に新規な配列のリスト。
【図12】GCF−IとGCF−II,配列認識番号1〜20、22〜29、32〜53及び169から由来のペプチド配列(後記の例2参照)を示し、特に新規な配列を示し、パネルAは、縮重オリゴヌクレオチドプローブと縮重PCRプライマーとを構成するのに使用するGCF−IIペプチドの配列がリストされ(配列認識番号45〜52)、パネルAのこれらの配列の内のいくつかは、合成ペプチドの構成に使用され、パネルBは、縮重プローブと縮重PCRプライマーの構成には短すぎた(6アミノ酸以下)新規なペプチドの配列のリスト(配列認識番号53)。
【図13】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図14】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図15】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図16】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図17】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図18】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図19】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図20】例3に関し、本発明の因子の細胞分裂誘発活性を示すグラフ。
【図21】例4に関し、図10のパネルA及び図12のパネルAに示された新規なペプチド配列から構成された縮重オリゴヌクレオチドのリスト(配列認識番号54〜88)。
【図22】例4に関し、縮重オリゴヌクレオチドプローブ609,650(それぞれ図21,配列認識番号69及び72参照)の結合サイトを含有する組替えウシゲノムファーゼGCF2BG1からの推定ゲノムファーゼGGF2BG1のストレッチを示すリストであって、同図は、DNA配列のコード化ストランドと第3読み取り枠の推定されたアミノ酸配列であり、因子2(太字)からのペプチド12の配列は、66アミノ酸転写読み取り枠(ヌクレオチド75272)の一部である。
【図23】例4に関し、後部下垂体からのRNAに存在するウシGGF−IIコード下配列のセグメントを分離する実験に使用された縮重PCRプライマー(パネルA,配列認識番号90〜108)とユニークなPCRプライマー(パネルB,配列認識番号109〜119)のリスト。
【図24】例4に関し、図7のパネルA,Bのプライマーのリストを使用したPCR増幅実験において得られた9つの別々の隣接したウシGGF−II cDNA構造体と配列、及び後部下垂体からのRNAを示し、同図の最上ラインは、特徴付けられたcDNA構造体に寄与するコード化配列の略図。
【図25】例4に関し、GGF2BG1のウシ組替えファーゼの物理的地図であり、前記ウシフラグメントは、長さが約20kbであり、ウシGGF−II遺伝子の二つのエクソン(太字)を有している。酵素Xbal,SpeI,Ndel,EcoRI,Kpnl,SstIの制限サイトがこの物理的地図上に示され、斜線部分は、配列のためにサブクローンされたフラグメントに対応する。
【図26】例4に関し、前記推定ウシGGF−II遺伝子の三つの別の遺伝子生成物の構造の略図であり、エクソンはその発見の順番にAないしEとしてリストされている。別のスプライシングパターン1,2及び3は、三つのオーバーラップする推定タンパク質構造体(GGF2BPP1,2,3)を生成し、これらはそれぞれ別の図28A,B,C(下記)に示されている。
【図27】配列認識番号120〜132を示し、図28A,28B,28C(下記)に示された推定タンパク質配列において同定されたGGF−I及びGGF−IIと、図10及び12にリストされた新規なペプチド配列との比較を示すリストであり、同図は、9つの新規なGGF−IIペプチド配列の内の6つが、これらの推定タンパク質配列に見られることを示しており、GGF−I配列に類似の二つのペプチド配列も見られる。
【図28A】、配列認識番号133を示し、図26のスプライシングパターンNo.1から得られるコード化トスランドDNAと推定アミノ酸配列のリスト(配列認識番号133)であり、前記推定ウシGGF−II遺伝子のこの部分cDNAは、206アミノ酸のタンパク質をコード化し、太字のペプチドは、図10及び12に示したリストから同定されたペプチドである。潜在的グリコシル化サイトがアンダーラインされている(ポリアデニル化信号AATAAAとともに)。
【図28B】配列認識番号134を示し、図26のスプライシングパターンNo.2から得られるコード化ストランドDNAと推定アミノ酸配列のリストであり、前記推定ウシGGF−II遺伝子のこの部分cDNAは、281アミノ酸のタンパク質をコード化し、太字のペプチドは、図10及び12に示したリストから同定されたペプチドである。潜在的グリコシル化サイトがアンダーラインされている(ポリアデニル化信号AATAAAとともに)。
【図28C】配列認識番号135を示し、図26のスプライシングパターンNo.3から得られるコード化トスランドDNAと推定アミノ酸配列のリストであり、前記推定ウシGGF−II遺伝子のこの部分cDNAは、257アミノ酸のタンパク質をコード化し、太字のペプチドは、図10及び12に示したリストから同定されたペプチドである。潜在的グリコシル化サイトがアンダーラインされている(ポリアデニル化信号AATAAAとともに)。
【図29】例6に関し、サザンブロット上の様々なほ乳類DNAに対する推定ウシGGF−II遺伝子配列のクロスハイブリダイゼーション分析のオートラジオグラムであり、前記フィルターは、同図にリストされた種からのEcoRI−消化DNA(各レーンにつき5μg)を含有する。前記プローブは、図25の物理的地図によって予測されるウシDNAの4キロベースのフラグメントを含んで、各DNAサンプルにおける単一の強力なバンドを検出し、より力の小さいバンドも観察され、これらは関連DNA配列を示す。他のほ乳類DNAサンプルからの強いハイブリダイゼーションバンドは、これらの種のGGF−II相同体を示すものと考えられる。
【図30】さまざまなスプライシング変異体を示す図であり、コード化セグメントは、F,E,B,A,G,C,C/D,C/D’,D,D’,H,K及びLによって示され、精製タンパク質からのペプチド配列の位置は、”0”によって示されている。
【図31】配列認識番号136〜147、160、161を示し、GGFのコード化セグメントのDNA配列と予想ペプチド配列のリストであり、ライン1は、ウシGGFの予想アミノ酸配列のリストであり、ライン2は、ウシGGFのヌクレオチド配列のリストであり、ライン3は、ヒトGGF(ヘレグリン)のヌクレオチド配列のリストであり(ヌクレオチドベースのマッチングは縦線にて示してある)、ライン4は、前記予想ウシ配列とは異なる場合におけるヒトGGF/heregulinの予想アミノ酸配列のリストであり、コード化セグメントE、A’及びKは、ウシ配列のみを表し、コード化セグメントD’は、ヒト(ヘレグリン)配列のみを表す。
【図32】配列認識番号148を示し、BPP5の予想GGF2アミノ酸配列とヌクレオチド配列のリストであり、上方ラインは、ヌクレオチド配列であり、下方ラインは、予想アミノ酸配列である。
【図33】配列認識番号149を示し、GGF2BPP2の予想アミノ酸配列とヌクレオチド配列のリストであり、上方ラインは、ヌクレオチド配列であり、下方ラインは、予想アミノ酸配列である。
【図34】配列認識番号150を示し、GGF2BPP4の予想アミノ酸配列とヌクレオチド配列のリストであり、上方ラインは、ヌクレオチド配列であり、下方ラインは、予想アミノ酸配列である。
【図35】配列認識番号151〜152を示し、二つのGGFペプチド配列(GGF2pp4とGGF2pp5)のヒトEGF(hEGF)との整合を示す配列のリストであり、星印は、保存されたシステインを示す。
【図36】GGFの増加に対する、約200kD(抗燐酸化ポリクローナル抗体とともに現像したウエスタンブロット上における200kDバンドの強度)のタンパク質のGGF活性のレベル(シュワン細胞細胞分裂誘発分析)と、チロシン燐酸化を示すグラフ。
【図37】図31の配列からのスプライシング変異体の配列のリスト。
【図38】EGFL1(配列認識番号154)の、予想アミノ酸配列を下部に、核酸配列を上部に示す配列のリスト。
【図39】EGFL2(配列認識番号155)の、予想アミノ酸配列を下部に、核酸配列を上部に示す配列のリスト。
【図40】EGFL3(配列認識番号156)の、予想アミノ酸配列を下部に、核酸配列を上部に示す配列のリスト。
【図41】EGFL4(配列認識番号157)の、予想アミノ酸配列を下部に、核酸配列を上部に示す配列のリスト。
【図42】EGFL5(配列認識番号158)の、予想アミノ酸配列を下部に、核酸配列を上部に示す配列のリスト。
【図43】EGFL6(配列認識番号159)の、予想アミノ酸配列を下部に、核酸配列を上部に示す配列のリスト。
【図44】クローンのスケールコード化セグメントマップであり、T3は、前記クローンからmRNAを生成するのに使用するバクテリオファージプロモーターを示し、R=隣接のEcoRI制限酵素サイトを、5’UTは、5’非翻訳領域を示し、E,B,A,C,C/D’及びDは、コード化セグメントを示し、0=翻訳開始サイト。ヒ=ウシEセグメントに対して相同の領域の5’限界(例6参照)、そして、3’UTは3’非翻訳領域を示す。
【図45】GGF2HBS5の予想アミノ酸配列(中間部:配列認識番号170)と、核酸配列(上部:配列認識番号21)とを示す。下部(中間)配列は、GGF−II調製物(図11及び12参照)からのペプチド配列を示す配列のリスト。
【図46】組替えヒト及びウシグリア成長因子のシュワン細胞細胞分裂誘発活性を示すグラフ。
【図47】CHO細胞のならし培地の大きさの異なる部分標本(aliquots)の投与から生じたシュワン細胞増殖活性物質の投与量−反応曲線。
【図48】GGF2HBS5cDNAクローンを含有するバキュロウィルスによって感染したSF9昆虫細胞によって細胞外培地中に分泌されたシュワン細胞細胞分裂誘発活性物質の投与量−反応曲線、
【図49】GGFペプチド抗体を使用した組替えCHO細胞のならし培地のウェスタンブロットを示す図。
【図50A】陽イオン交換カラムから溶出した組替え(COS細胞が生成)ヒトGGF−II(rhGGF−II)ピークのシュワン細胞増殖活性のグラフ。
【図50B】rhGGFIIの特定のペプチドに対して生成されたポリクローナル抗体を使用した組替えGGFIIピークに対する免疫ブロット
【図51A】フラクションごとの陽イオン交換カラムでのrhGGF−II(CHO細胞が生成)の精製を示すグラフ。
【図51B】図51Aに示されたフラクションとrhGGF−II特異抗体を使用したウエスタンブロットの写真。
【図52】組替えグリア成長因子で処理したシュワン細胞におけるチロシン燐酸化を示すゲルの写真。
【図53】GGFHBS5,GGFHFB1及びGGFBPP5ポリペプチド(配列認識番号170,171,172)の配列のリスト。
【図54】前記CHO細胞−発現ベクターpcDHFRpolyAのマップ。
【配列表】
Claims (9)
- GGF/p185 erbB2 リガンド遺伝子によってコードされるポリペプチドであって、
p185 erbB2 レセプターと相互作用し、かつ配列認識番号170のアミノ酸配列を含むポリペプチド。 - 前記ポリペプチドが、グリア細胞の細胞分裂誘発を誘発する請求項1に記載のポリペプチド。
- 請求項1に記載のポリペプチドをコードする単離核酸分子。
- 前記単離核酸分子が、配列認識番号21の塩基配列を含む請求項3に記載の単離核酸分子。
- プロモーターに作動連結された請求項3に記載の単離核酸分子を含む組換えベクター。
- 請求項3に記載の単離核酸分子でトランスフォーム、またはトランスフェクトされた細胞ライン。
- 請求項1に記載のポリペプチドを生体外で産生する方法であって、
請求項6に記載の細胞ラインを提供する工程、前記細胞ラインを前記核酸分子の発現を許容する条件下で培養する工程、そして前記ポリペプチドを単離する工程、を有する方法。 - サンプル中における請求項1に記載のポリペプチドに対するレセプターの存在を同定する方法であって、
前記サンプルを前記ポリペプチドに接触させる工程、及びそれの間の結合を測定する工程を有し、前記結合が前記レセプターの存在の指標である、方法。 - 許容可能な希釈剤、担体または賦形剤、および/または単位製剤での、医薬または獣医薬用に調製された請求項1に記載のポリペプチドを有する医薬または獣医薬用製剤。
Applications Claiming Priority (9)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US90713892A | 1992-06-30 | 1992-06-30 | |
| US07/907,138 | 1992-06-30 | ||
| US94038992A | 1992-09-03 | 1992-09-03 | |
| US07/940,389 | 1992-09-03 | ||
| US96517392A | 1992-10-23 | 1992-10-23 | |
| US07/965,173 | 1992-10-23 | ||
| US08/036,555 US5530109A (en) | 1991-04-10 | 1993-03-24 | DNA encoding glial mitogenic factors |
| US08/036,555 | 1993-03-24 | ||
| PCT/US1993/006228 WO1994000140A1 (en) | 1992-06-30 | 1993-06-29 | Glial mitogenic factors, their preparation and use |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPH08509357A JPH08509357A (ja) | 1996-10-08 |
| JP4127567B2 true JP4127567B2 (ja) | 2008-07-30 |
Family
ID=27488375
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP50265594A Expired - Lifetime JP4127567B2 (ja) | 1992-06-30 | 1993-06-29 | グリア細胞分裂誘発因子とその調製及び使用 |
Country Status (11)
| Country | Link |
|---|---|
| US (8) | US5530109A (ja) |
| EP (1) | EP0653940B1 (ja) |
| JP (1) | JP4127567B2 (ja) |
| KR (2) | KR100307943B1 (ja) |
| AT (1) | ATE286126T1 (ja) |
| AU (2) | AU688270B2 (ja) |
| CA (1) | CA2139136A1 (ja) |
| DE (1) | DE69333731T2 (ja) |
| ES (1) | ES2236686T3 (ja) |
| PT (1) | PT101297B (ja) |
| WO (1) | WO1994000140A1 (ja) |
Families Citing this family (78)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20050106666A1 (en) * | 1991-04-10 | 2005-05-19 | Cambridge Neuroscience | Glial mitogenic factors, their preparation and use |
| US5716930A (en) * | 1991-04-10 | 1998-02-10 | Ludwig Institute For Cancer Research | Glial growth factors |
| US5530109A (en) * | 1991-04-10 | 1996-06-25 | Ludwig Institute For Cancer Research | DNA encoding glial mitogenic factors |
| US7319019B1 (en) * | 1991-04-10 | 2008-01-15 | Acorda Therapeutics, Inc. | Glial mitogenic factors lacking an N-terminal signal sequence |
| US7115554B1 (en) * | 1993-05-06 | 2006-10-03 | Acorda Therapeutics, Inc. | Methods of increasing myotube formation or survival or muscle cell mitogenesis differentiation or survival using neuregulin GGF III |
| US7037888B1 (en) * | 1992-04-03 | 2006-05-02 | Acorda Therapeutics, Inc. | Methods for treating muscle diseases and disorders |
| US6087323A (en) * | 1992-04-03 | 2000-07-11 | Cambridge Neuroscience, Inc. | Use of neuregulins as modulators of cellular communication |
| AU693767B2 (en) * | 1992-08-10 | 1998-07-09 | Cambridge Neuroscience, Inc. | Inhibitors of cell proliferation, their preparation and use |
| US5770567A (en) * | 1994-11-14 | 1998-06-23 | Genentech, Inc. | Sensory and motor neuron derived factor (SMDF) |
| US6750196B1 (en) * | 1995-03-27 | 2004-06-15 | Acorda Therapeutics | Methods of treating disorders of the eye |
| US5670342A (en) * | 1995-04-06 | 1997-09-23 | Amgen Inc. | NDF peptides |
| US6033660A (en) * | 1995-05-10 | 2000-03-07 | Genentech, Inc. | Method of treating a nervous system injury with cultured schwann cells |
| US5721139A (en) * | 1995-05-10 | 1998-02-24 | Genentech, Inc. | Isolating and culturing schwann cells |
| US5849585A (en) * | 1995-05-10 | 1998-12-15 | Genetech, Inc. | Isolating and culturing Schwann cells |
| GB9514325D0 (en) | 1995-07-13 | 1995-09-13 | Royal Free Hosp School Med | Detection and quantification of human herpes virus 7 by enzymic amplification |
| US5912326A (en) * | 1995-09-08 | 1999-06-15 | President And Fellows Of Harvard College | Cerebellum-derived growth factors |
| WO1998002540A1 (en) | 1996-07-12 | 1998-01-22 | Genentech, Inc. | Chimeric heteromultimer adhesins |
| ATE290595T1 (de) | 1996-07-12 | 2005-03-15 | Genentech Inc | Gamma-heregulin |
| US5955111A (en) * | 1996-10-08 | 1999-09-21 | Hartford Hospital | Methods and compositions for inducing production of stress proteins |
| US6136558A (en) * | 1997-02-10 | 2000-10-24 | Genentech, Inc. | Heregulin variants |
| ATE427353T1 (de) | 1997-02-10 | 2009-04-15 | Genentech Inc | Heregulin varianten |
| CA2306228A1 (en) * | 1997-10-14 | 1999-04-22 | Cambridge Neuroscience, Inc. | Therapeutic methods comprising use of a neuregulin |
| WO1999031505A1 (en) * | 1997-12-15 | 1999-06-24 | Shire Laboratories Inc. | Neurotrophic factor assay |
| US20010023241A1 (en) | 1998-02-04 | 2001-09-20 | Sliwkowski Mark X. | Use of heregulin as a growth factor |
| US6110718A (en) * | 1998-03-20 | 2000-08-29 | Wayne State University | Mammalian putative phosphatidylinositol-4-phosphate-5-kinase |
| US6051401A (en) * | 1998-07-28 | 2000-04-18 | Bayer Corporation | Methods and constructs for protein expression |
| AUPP785098A0 (en) * | 1998-12-21 | 1999-01-21 | Victor Chang Cardiac Research Institute, The | Treatment of heart disease |
| US6635249B1 (en) | 1999-04-23 | 2003-10-21 | Cenes Pharmaceuticals, Inc. | Methods for treating congestive heart failure |
| US6933122B1 (en) * | 1999-05-14 | 2005-08-23 | The Trustees Of Columbia University In The City Of New York | A-form of cytoplasmic domain of nARIA (CRD-neuregulin) and uses thereof |
| ES2252216T3 (es) * | 2000-02-11 | 2006-05-16 | Proteosys Ag | Utilizacion de neorregulina-beta como indicador y/o diana. |
| EP1681299B1 (en) * | 2000-03-27 | 2009-11-11 | Genetics Institute, LLC | Methods for purifying highly anionic proteins |
| ES2370875T3 (es) | 2000-05-23 | 2011-12-23 | Acorda Therapeutics, Inc. | Moléculas de ácido nucleico nrg-2, polipéptidos y métodos de diagnóstico y terapéuticos. |
| EP1417230A2 (en) * | 2001-08-06 | 2004-05-12 | ProteoSys AG | Neuregulin-beta isoforms associated with neuronal processes |
| AU2002304965A1 (en) * | 2002-05-24 | 2003-12-12 | Zensun (Shanghai) Sci-Tech.Ltd | Neuregulin based methods and compositions for treating viral myocarditis and dilated cardiomyopathy |
| CA2536146A1 (en) * | 2003-08-19 | 2005-02-24 | Agos Biotech Ltd. | Splice variants of erbb ligands, compositions and uses thereof |
| TWI276685B (en) * | 2003-12-30 | 2007-03-21 | Ind Tech Res Inst | Conditional medium for culturing Schwann cells |
| EP1781709A4 (en) * | 2004-08-27 | 2011-07-06 | Absorbent Technologies Inc | SUPER ABSORBING POLYMERS IN AGRICULTURAL APPLICATIONS |
| KR100680479B1 (ko) * | 2005-04-11 | 2007-02-08 | 주식회사 하이닉스반도체 | 비휘발성 메모리 장치의 프로그램 검증 방법 |
| US20070213264A1 (en) | 2005-12-02 | 2007-09-13 | Mingdong Zhou | Neuregulin variants and methods of screening and using thereof |
| US20070190127A1 (en) | 2005-12-30 | 2007-08-16 | Mingdong Zhou | Extended release of neuregulin for improved cardiac function |
| US9580515B2 (en) | 2006-08-21 | 2017-02-28 | Zensun (Shanghai) Science & Technology, Co., Ltd. | Neukinase, a downstream protein of neuregulin |
| US7521535B2 (en) * | 2006-09-20 | 2009-04-21 | The Board Of Regents For Oklahoma State University | Anti-microbial defensin-related peptides and methods of use |
| CA2686959C (en) | 2007-05-10 | 2017-06-13 | Acorda Therapeutics Inc. | Methods for detecting cardiac damage |
| US20090156488A1 (en) * | 2007-09-12 | 2009-06-18 | Zensun (Shanghai) Science & Technology Limited | Use of neuregulin for organ preservation |
| AU2008323169B2 (en) | 2007-11-16 | 2013-08-29 | Mind-Nrg Sa. | Active soluble post-translationally modified neuregulin isoforms |
| EP2262527B1 (en) | 2008-02-29 | 2016-08-03 | Acorda Therapeutics, Inc. | Compositions for achieving desired glial growth factor 2 plasma levels |
| US9348975B2 (en) * | 2008-05-02 | 2016-05-24 | Baxter International Inc. | Optimizing therapy outcomes for peritoneal dialysis |
| US8882700B2 (en) | 2008-05-02 | 2014-11-11 | Baxter International Inc. | Smart patient transfer set for peritoneal dialysis |
| US8057679B2 (en) | 2008-07-09 | 2011-11-15 | Baxter International Inc. | Dialysis system having trending and alert generation |
| EP2320933B1 (en) | 2008-07-17 | 2017-12-27 | Acorda Therapeutics, Inc. | Therapeutic dosing of a neuregulin or a subsequence thereof for treatment or prophylaxis of heart failure |
| CN104707130A (zh) | 2008-08-15 | 2015-06-17 | 阿索尔达治疗公司 | 在cns神经性损伤后非-急性期间处理的组合物及方法 |
| JP5743898B2 (ja) | 2008-11-28 | 2015-07-01 | ゼンサン (シャンハイ) サイエンス アンド テクノロジー リミテッド | ニューレグリンペプチド及びその使用 |
| JP2012509908A (ja) * | 2008-11-28 | 2012-04-26 | ゼンサン (シャンハイ) サイエンス アンド テクノロジー リミテッド | ニューレグリン及び心臓幹細胞 |
| US8377012B2 (en) * | 2009-01-30 | 2013-02-19 | Baxter International Inc. | Transfer sets for therapy optimization |
| US8282829B2 (en) | 2009-05-20 | 2012-10-09 | Baxter International Inc. | System and method for automated data collection of twenty-four hour ultrafiltration and other patient parameters using wired or wireless technology |
| ES2748886T3 (es) | 2009-06-09 | 2020-03-18 | Zensun Shanghai Science & Tech Co Ltd | Métodos basados en Neuregulina para el tratamiento de la insuficiencia cardíaca |
| EP3087998B1 (en) | 2009-06-09 | 2019-09-04 | Zensun (Shanghai) Science & Technology, Co., Ltd. | Neuregulin based methods for treating heart failure |
| JP6096262B2 (ja) | 2009-08-25 | 2017-03-15 | ゼンサン (シャンハイ) サイエンス アンド テクノロジー,シーオー.,エルティーディー. | ニューレグリンに基づく心不全の治療方法 |
| NZ625673A (en) | 2009-10-14 | 2015-11-27 | Acorda Therapeutics Inc | Use of a neuregulin to treat peripheral nerve injury |
| US9020827B2 (en) * | 2009-10-16 | 2015-04-28 | Baxter International Inc. | Peritoneal dialysis optimized using a patient hand-held scanning device |
| CN102139095A (zh) | 2010-01-29 | 2011-08-03 | 上海泽生科技开发有限公司 | 神经调节蛋白用于预防、治疗或延迟心脏缺血再灌注损伤的方法和组合物 |
| WO2013053076A1 (en) | 2011-10-10 | 2013-04-18 | Zensun (Shanghai)Science & Technology Limited | Compositions and methods for treating heart failure |
| DK2830645T3 (da) | 2012-03-30 | 2017-11-20 | Acorda Therapeutics Inc | Neuregulin til anvendelse i behandling af perifær nervelæsion |
| RU2650635C2 (ru) | 2012-10-08 | 2018-04-16 | Цзэньсунь (Шанхай) Сайенс Энд Текнолоджи, Ко., Лтд. | Композиции и способы лечения сердечной недостаточности у пациентов с диабетом |
| CA2904055A1 (en) | 2013-03-06 | 2014-09-12 | Acorda Therapeutics, Inc. | Therapeutic dosing of a neuregulin or a fragment thereof for treatment or prophylaxis of heart failure |
| CN105611955A (zh) | 2013-05-22 | 2016-05-25 | 上海泽生科技开发有限公司 | 缓释纽兰格林治疗心力衰竭 |
| CN104758922A (zh) | 2014-01-03 | 2015-07-08 | 上海泽生科技开发有限公司 | 纽兰格林制剂的配方 |
| WO2015140351A1 (en) | 2014-03-21 | 2015-09-24 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Methods and pharmaceutical compositions for enhancing myelination |
| CN105497876B (zh) | 2014-09-24 | 2021-01-15 | 上海泽生科技开发股份有限公司 | 神经调节蛋白用于预防、治疗或延迟心脏室性心律失常的方法和组合物 |
| CN105561298A (zh) | 2014-10-17 | 2016-05-11 | 上海泽生科技开发有限公司 | 神经调节蛋白用于预防、治疗或延迟射血分数保留的心力衰竭的方法和组合物 |
| EP3353546A1 (en) | 2015-09-25 | 2018-08-01 | Douglas B. Sawyer | Methods for treating cardiac injury |
| WO2019200033A1 (en) | 2018-04-11 | 2019-10-17 | Salubris Biotherapeutics, Inc. | Human neuregulin-1 (nrg-1) recombinant fusion protein compositions and methods of use thereof |
| CN111407881A (zh) | 2019-01-07 | 2020-07-14 | 上海泽生科技开发股份有限公司 | 神经调节蛋白用于预防、治疗或延迟心肌损伤的方法和组合物 |
| AR121035A1 (es) | 2019-04-01 | 2022-04-13 | Lilly Co Eli | Compuestos de neuregulina-4 y métodos de uso |
| US20230057622A1 (en) | 2019-09-16 | 2023-02-23 | Zensun (Shanghai) Science & Technology, Co., Ltd. | Recombinant Human Neuregulin Derivatives and Use Thereof |
| CN113289002A (zh) | 2020-02-24 | 2021-08-24 | 上海泽生科技开发股份有限公司 | 神经调节蛋白用于预防、治疗或延缓心力衰竭的方法和组合物 |
| EP4351722A1 (en) | 2021-06-10 | 2024-04-17 | Amgen Inc. | Engineered nrg-1 variants with improved selectivity toward erbb4 but not against erbb3 |
| WO2023178086A1 (en) | 2022-03-15 | 2023-09-21 | Salubris Biotherapeutics, Inc. | Methods of treating fibrosis and arrhythmia with a neuregulin-1 fusion protein |
Family Cites Families (19)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4935341A (en) * | 1986-06-04 | 1990-06-19 | Whitehead Institute For Biomedical Research | Detection of point mutations in neu genes |
| US5214034A (en) * | 1986-09-26 | 1993-05-25 | Mitsui Toatsu Chemicals, Incorporated | Catechol derivatives, and preventive and remedial preparations for regressive disorders in the central nervous system containing the same |
| US5019559A (en) * | 1986-11-14 | 1991-05-28 | President And Fellows Of Harvard College | Wound healing using PDGF and IGF-II |
| US4968603A (en) * | 1986-12-31 | 1990-11-06 | The Regents Of The University Of California | Determination of status in neoplastic disease |
| WO1989006692A1 (en) * | 1988-01-12 | 1989-07-27 | Genentech, Inc. | Method of treating tumor cells by inhibiting growth factor receptor function |
| US5057494A (en) * | 1988-08-03 | 1991-10-15 | Ethicon, Inc. | Method for preventing tissue damage after an ischemic episode |
| WO1990014357A1 (en) * | 1989-05-19 | 1990-11-29 | Genentech, Inc. | Her2 extracellular domain |
| US5235043A (en) * | 1990-04-06 | 1993-08-10 | Synergen, Inc. | Production of biologically active, recombinant members of the ngf/bdnf family of neurotrophic proteins |
| WO1991015230A1 (en) * | 1990-04-06 | 1991-10-17 | The Trustees Of The University Of Pennsylvania | Ligand for the neu gene product |
| AU667026B2 (en) * | 1990-05-25 | 1996-03-07 | Georgetown University | Growth factor which inhibits the growth of cells overexpressing the human oncogene erbB-2 |
| US5202428A (en) * | 1990-06-20 | 1993-04-13 | The Salk Institute For Biological Studies | DNA encoding neurotropic growth factor |
| ATE182152T1 (de) * | 1991-01-14 | 1999-07-15 | Univ Georgetown | Wachstumsfaktor-ligand, der an den erbb-2- rezeptor bindet und die zellulären reaktionen induziert |
| GB9107566D0 (en) * | 1991-04-10 | 1991-05-29 | Ludwig Inst Cancer Res | Glial mitogenic factors,their preparation and use |
| US5530109A (en) * | 1991-04-10 | 1996-06-25 | Ludwig Institute For Cancer Research | DNA encoding glial mitogenic factors |
| US5367060A (en) * | 1991-05-24 | 1994-11-22 | Genentech, Inc. | Structure, production and use of heregulin |
| IL101943A0 (en) * | 1991-05-24 | 1992-12-30 | Genentech Inc | Structure,production and use of heregulin |
| US5237056A (en) * | 1991-05-29 | 1993-08-17 | President And Fellows Of Harvard College | DNA encoding a protein which copurifies with acetylcholine receptor inducing activity and uses therefor |
| WO1993022424A1 (en) * | 1992-04-29 | 1993-11-11 | Amgen Inc. | RECOMBINANT STIMULATING FACTOR OF THE neu RECEPTOR |
| JPH08502241A (ja) * | 1992-04-29 | 1996-03-12 | ジョージタウン・ユニバーシティ | 上皮増殖因子受容体およびerbB−2受容体のリガンド増殖因子と相互作用する結合ペプチド |
-
1993
- 1993-03-24 US US08/036,555 patent/US5530109A/en not_active Expired - Lifetime
- 1993-06-29 WO PCT/US1993/006228 patent/WO1994000140A1/en active IP Right Grant
- 1993-06-29 AT AT93918139T patent/ATE286126T1/de not_active IP Right Cessation
- 1993-06-29 DE DE69333731T patent/DE69333731T2/de not_active Expired - Lifetime
- 1993-06-29 AU AU47694/93A patent/AU688270B2/en not_active Expired
- 1993-06-29 ES ES93918139T patent/ES2236686T3/es not_active Expired - Lifetime
- 1993-06-29 EP EP93918139A patent/EP0653940B1/en not_active Expired - Lifetime
- 1993-06-29 KR KR1019940704870A patent/KR100307943B1/ko not_active IP Right Cessation
- 1993-06-29 KR KR1020007004157A patent/KR100344006B1/ko not_active IP Right Cessation
- 1993-06-29 CA CA002139136A patent/CA2139136A1/en not_active Abandoned
- 1993-06-29 JP JP50265594A patent/JP4127567B2/ja not_active Expired - Lifetime
- 1993-06-30 PT PT101297A patent/PT101297B/pt not_active IP Right Cessation
-
1995
- 1995-06-06 US US08/471,855 patent/US5621081A/en not_active Expired - Lifetime
- 1995-06-06 US US08/472,008 patent/US5602096A/en not_active Expired - Lifetime
- 1995-06-06 US US08/470,335 patent/US6147190A/en not_active Expired - Lifetime
- 1995-06-06 US US08/469,526 patent/US5792849A/en not_active Expired - Lifetime
- 1995-06-06 US US08/469,569 patent/US5606032A/en not_active Expired - Lifetime
- 1995-06-06 US US08/470,339 patent/US6232286B1/en not_active Expired - Lifetime
-
1996
- 1996-10-22 US US08/734,591 patent/US5854220A/en not_active Expired - Lifetime
-
1997
- 1997-12-12 AU AU48338/97A patent/AU4833897A/en not_active Withdrawn
Also Published As
| Publication number | Publication date |
|---|---|
| PT101297B (pt) | 1999-10-29 |
| EP0653940A1 (en) | 1995-05-24 |
| KR100307943B1 (ko) | 2001-12-01 |
| US5530109A (en) | 1996-06-25 |
| EP0653940A4 (en) | 1997-02-26 |
| PT101297A (pt) | 1994-06-30 |
| AU4769493A (en) | 1994-01-24 |
| US5606032A (en) | 1997-02-25 |
| US5792849A (en) | 1998-08-11 |
| AU4833897A (en) | 1998-04-23 |
| AU688270B2 (en) | 1998-03-12 |
| KR100344006B1 (ko) | 2002-12-11 |
| US5854220A (en) | 1998-12-29 |
| DE69333731D1 (de) | 2005-02-03 |
| US5602096A (en) | 1997-02-11 |
| ATE286126T1 (de) | 2005-01-15 |
| DE69333731T2 (de) | 2006-03-09 |
| US6232286B1 (en) | 2001-05-15 |
| EP0653940B1 (en) | 2004-12-29 |
| US6147190A (en) | 2000-11-14 |
| US5621081A (en) | 1997-04-15 |
| JPH08509357A (ja) | 1996-10-08 |
| ES2236686T3 (es) | 2005-07-16 |
| WO1994000140A1 (en) | 1994-01-06 |
| CA2139136A1 (en) | 1994-01-06 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4127567B2 (ja) | グリア細胞分裂誘発因子とその調製及び使用 | |
| US5716930A (en) | Glial growth factors | |
| KR100284909B1 (ko) | 근육 질병 및 장애 치료용 조성물 | |
| US7968678B2 (en) | Glial mitogenic factors, their preparation and use | |
| US5876973A (en) | Method for making antibodies which specifically bind to glial growth factors | |
| US7541338B2 (en) | Glial mitogenic factors, their preparation and use | |
| US7135456B1 (en) | Glial mitogenic factors, their preparation and use | |
| US7094749B1 (en) | Glial mitogenic factors, their preparation and use | |
| US20050106666A1 (en) | Glial mitogenic factors, their preparation and use |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20060718 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20061017 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20061204 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070117 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20070327 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20070626 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20070813 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20070726 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20070910 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20070824 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20071001 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070927 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20071120 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20080319 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20080410 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20080424 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20080509 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110523 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110523 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120523 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130523 Year of fee payment: 5 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |