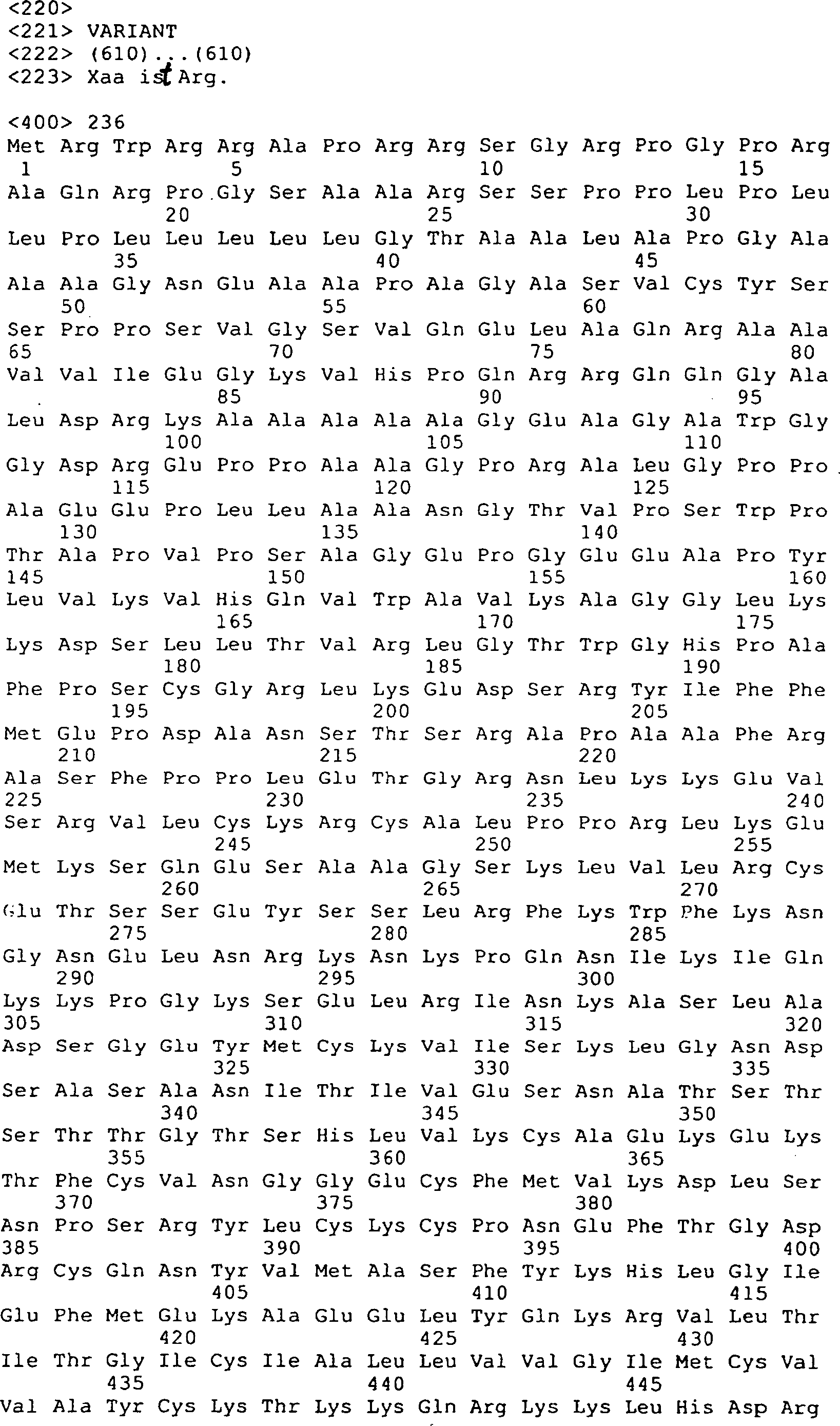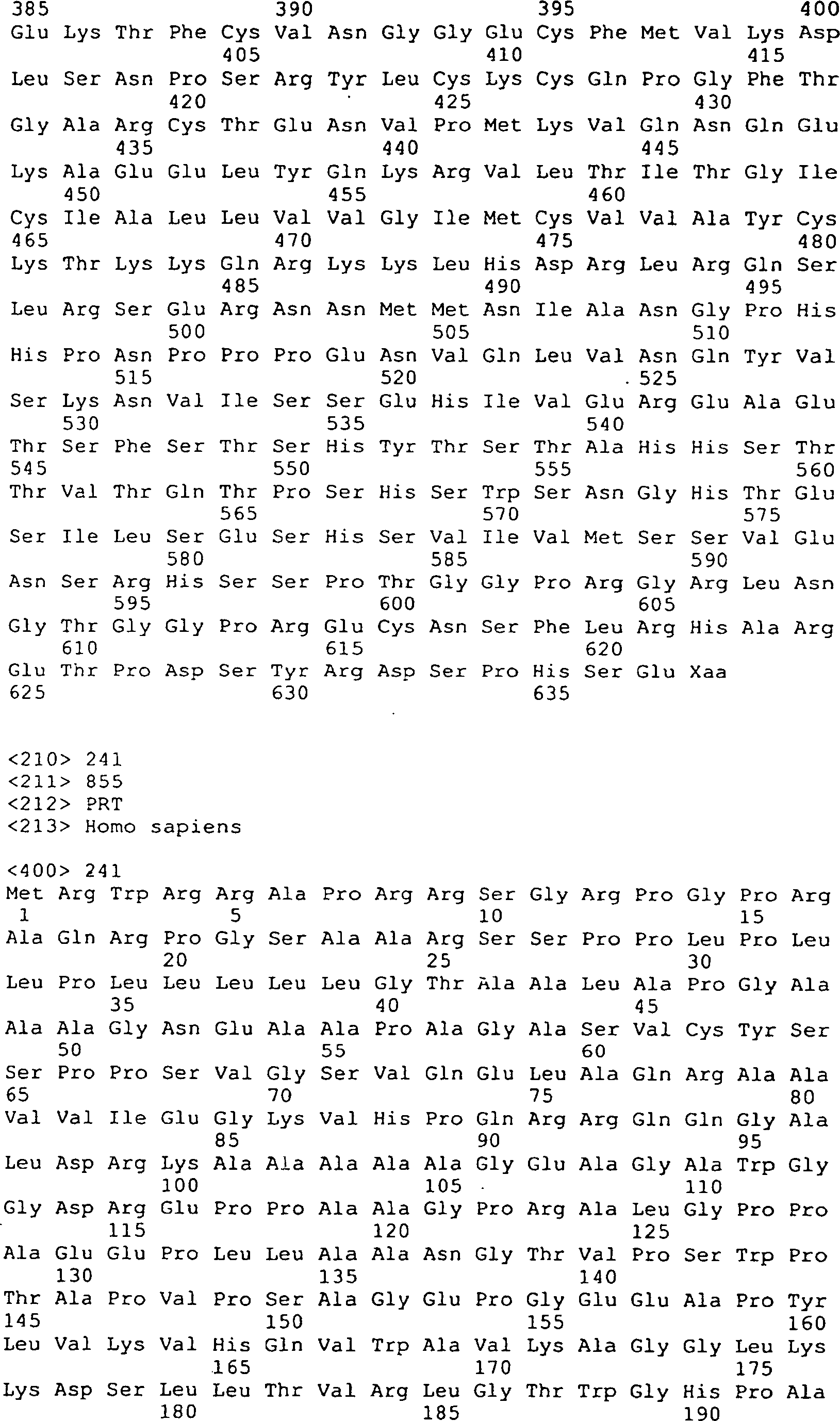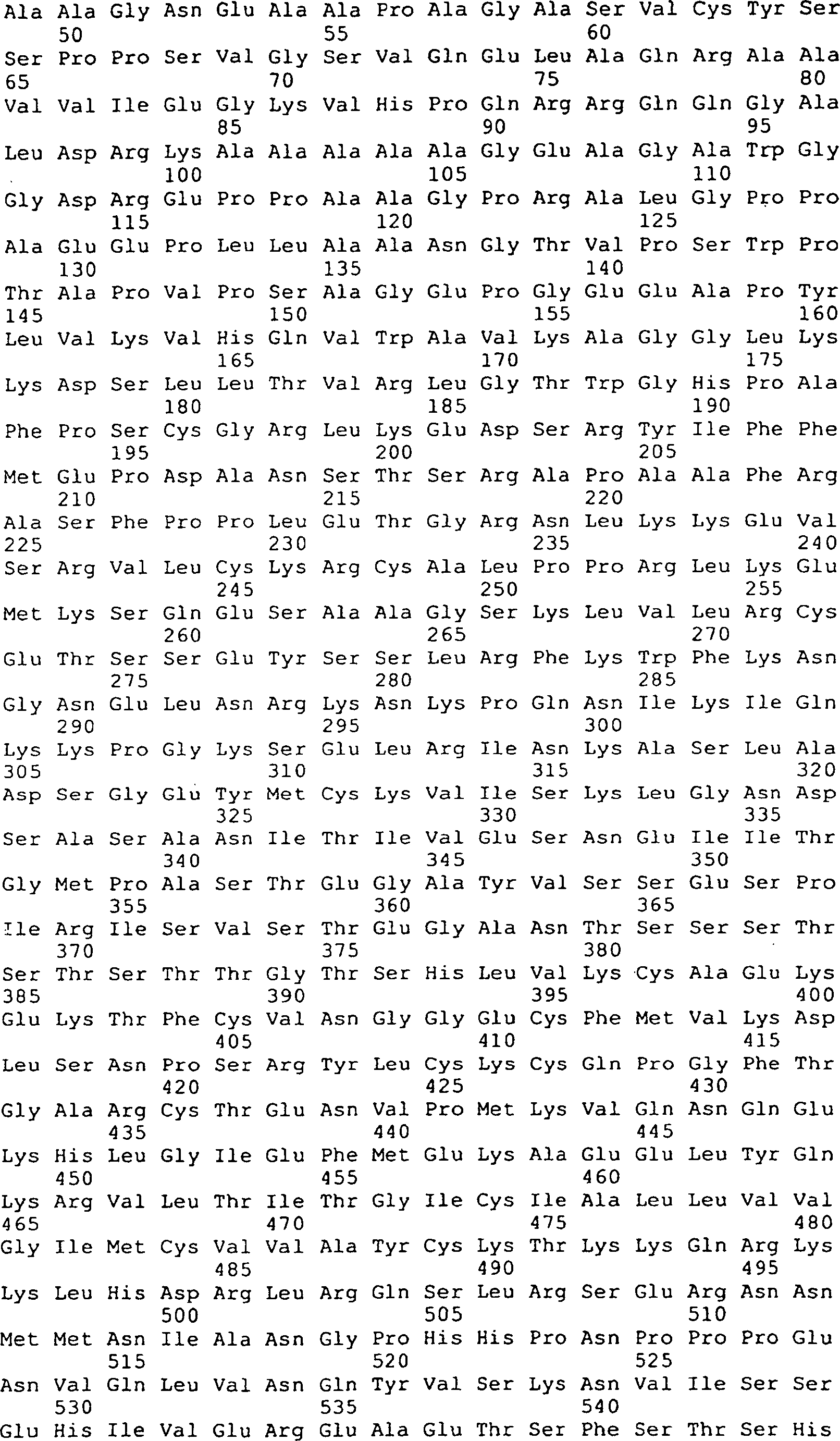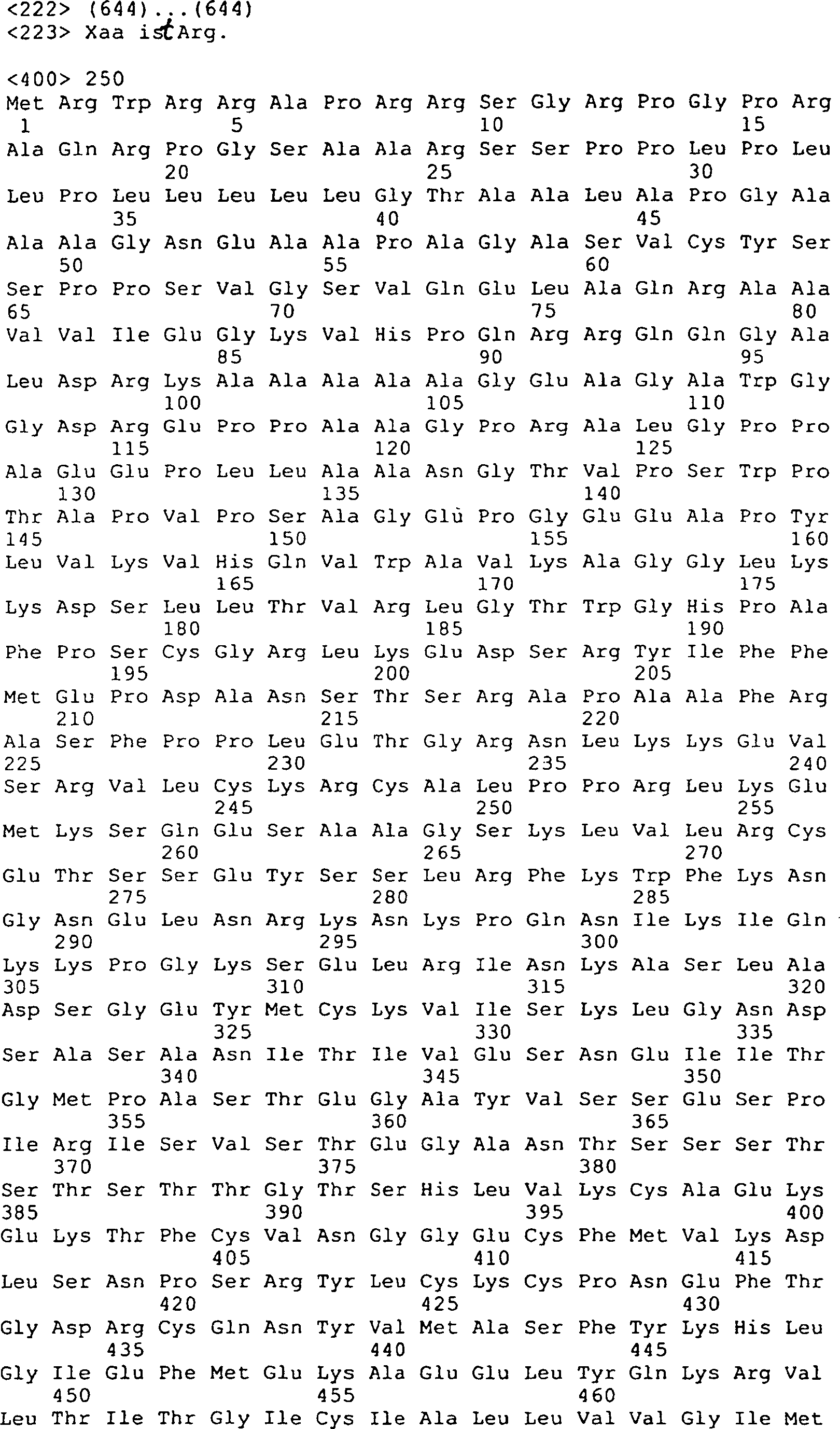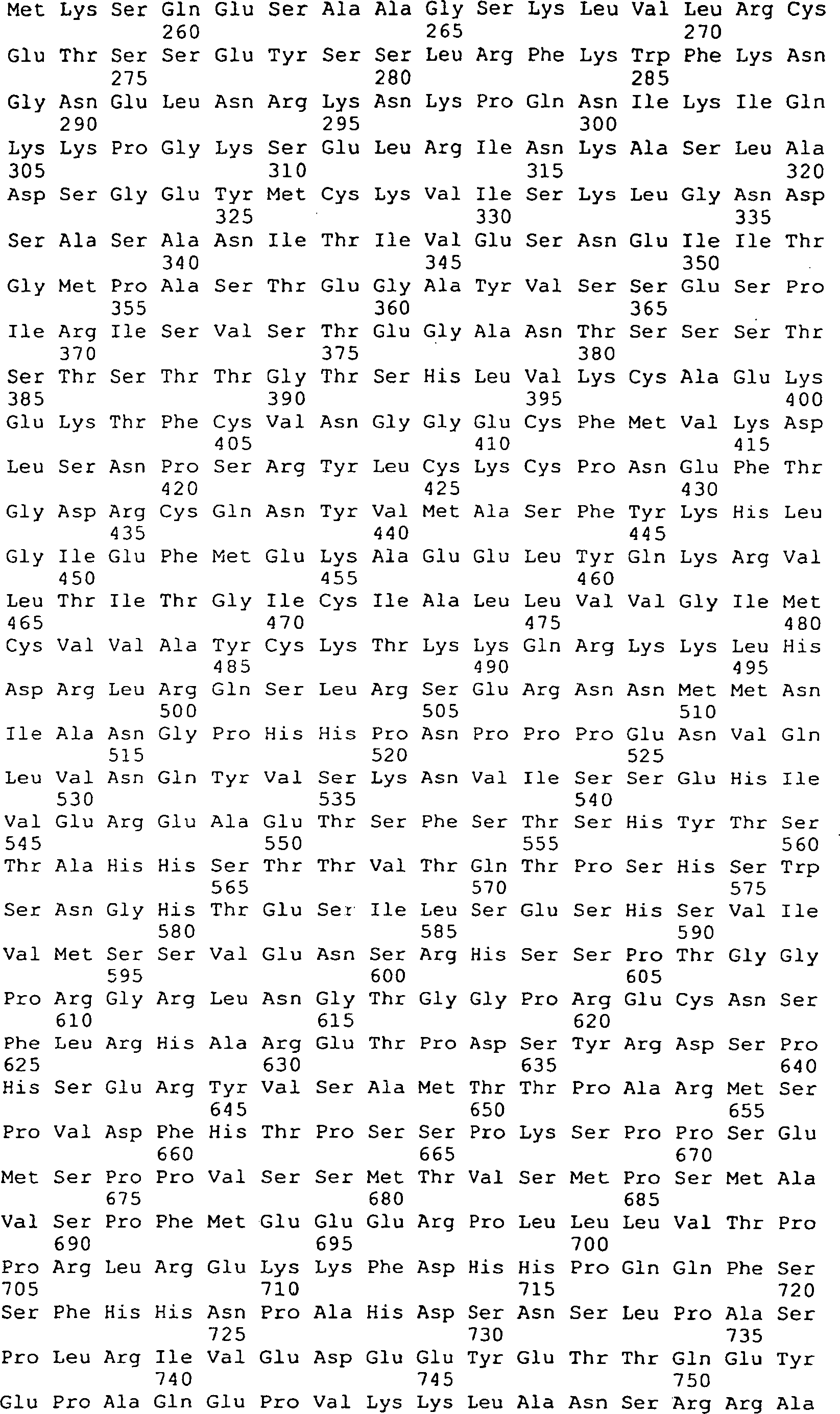-
Die
Erfindung bezieht sich auf in Wirbeltierarten gefundener Polypeptide,
die mitogenetische Wachstumsfaktoren für Gliazellen, einschließlich Schwann-Zellen, darstelle.
Die Erfindung betrifft ferner Verfahren zur Herstellung von solchen
Faktoren sowie die therapeutische Anwendung von diesen Faktoren.
-
Die
Gliazellen von Wirbeltieren stellen das spezialisierte Bindegewebe
des zentralen und peripheren Nervensystems dar. Wichtige Gliazellen
umfassen die Schwann-Zellen, die eine metabolische Unterstützung für die Neuronen
sowie die Myelinscheiden um die Axone von bestimmten peripheren
Neuronen, wodurch individuelle Nervenfasern gebildet werden, bereitstellen.
Die Schwann-Zellen unterstützen
die Neuronen und stellen eine Scheidenwirkung durch die Ausbildung
von konzentrischen Membranschichten um die benachbarten Nervenaxone
bereit, wobei sie sich bei ihrer Entwicklung um die Axone drehen.
Diese Myelinscheiden sind ein empfindliches Element von vielen Nervenfasern
und eine Beschädigung
der Schwann-Zellen oder ein Fehler im Wachstum und in der Entwicklung
kann mit einer signifikanten Demyelinisation oder Nervendegeneration assoziiert
sein, was für
eine Anzahl von Krankheiten und Störungen des peripheren Nervensystems
charakteristisch ist. Es wurde erkannt, dass bei der Entwicklung
des Nervensystems, die Zellen verschiedene Faktoren benötigen, um
ihre Teilung und ihr Wachstum zu steuern. Verschiedene Faktoren
wurden in den vergangenen Jahren identifiziert, wobei auch einige
gefunden wurden, die eine Wirkung auf die Teilung und die Entwicklung der
Schwann-Zellen besitzen.
-
Brockes
et al. beschreiben unter anderem in J. Neuroscience, 4 (1984), 75–83, einen
Proteinwachstumsfaktor, der in Extrakten vom Rindergehirn und Hypophysengewebe
vorhanden ist und der bezeichnet wurde als „Glial Growth Factor" (GGF). Dieser Faktor
stimuliert kultivierte Schwann-Zellen aus der Ratte zur Teilung
in einem Grundmedium, das 10% fötales
Kälberserum
enthielt. Es wurde auch beschrieben, dass dieser Faktor eine Molekülmasse von
31.000 Daltons hat und vollkommen dimerisiert ist. In Meth. Enz.,
147 (1987), 217–225,
beschreibt Brockes einen Test für
GGF mit Schwann-Zellen.
-
Brockes
et al., siehe oben, beschreiben auch ein Verfahren für die Reinigung
von GGF bis zur augenscheinlichen Homogenität. In Kürze umfasst ein beschriebenes,
größeres Reinigungsverfahren
die Extraktion aus lyophilisierten Vorderlappen vom Rind und die
Chromatographie des erzielten Materials unter Verwendung der NaCl-Gradientenelution
von CM-Cellulose. Die Gelfiltration wird dann mit einer Ultrogel-Säule durchgeführt, gefolgt
von einer Elution aus einer Phosphocellulosesäule. Abschließend erfolgt
eine SDS-Gelelektrophorese
im Labormaßstab.
Alternativ wird das CM-Cellulosematerial direkt auf eine Phosphorcellulosesäule aufgetragen,
die Fraktionen aus der Säule
werden vereinigt und mit einer präparativen, nativen Gelelektrophorese
und anschließend
durch eine abschließende
SDS-Gelelektrophorese gereinigt.
-
Brockes
et al. haben beobachtet, dass bei den zuvor berichteten Gelfiltrationsexperimenten
(Brockes et al., J. Biol. Chem. 255 (1980), 8374–8377) der Hauptpeak der Wachstumsfaktoraktivität bei einer
relativen Molekülmasse
von 56.000 Daltons auftrat, während
bei dem ersten der oben beschriebenen Verfahren die Aktivität vorwiegend
bei einer relativen Molekülmasse
von 31.000 beobachtet wurde. Es wurde berichtet, dass das GGF-Dimer
in einem großen
Umfang als Ergebnis der Gradientenelution aus der CM-Cellutose bei
diesem Verfahren entfernt wird.
-
Benveniste
et al. (PNAS, 82 (1985), 3930–3934)
beschreiben einen von T-Lymphozyten
abstammenden, das Gliawachstum fördernden
Faktor. Dieser Faktor zeigt unter reduzierenden Bedingungen einen
Wechsel in der relativen Molekülmasse
auf SDS-Gelen.
-
Kimura
et al. (Nature, 348 (1990), 257–260)
beschreiben einen Faktor, den sie als „Schwannoma-derived growth
factor" (SDGF) bezeichnet
haben, der von einem Tumor der Ischiasnervenscheiden abstammte. Die
Autoren haben vermerkt, dass SDGF nicht die Einlagerung von Tritium
markierten TdR in kultivierte Schwann-Zellen unter Bedingungen stimuliert,
wo eine partiell gereinigte Hypophysenfraktion, die GGF enthält, aktiv
ist. SDGF hat ein scheinbares Molekulargewicht zwischen 31.000 und
35.000.
-
Davis
und Stroobant (J. Cell. Biol., 110 (1990), 1353–1360) beschreiben die Testung
von einer Reihe von möglichen
Mitogenen. Es wurden Schwann-Zellen
aus der Ratte verwendet und die ausgewählten Kandidatensubstanzen
wurden hinsichtlich ihrer Fähigkeit überprüft, die
DNA-Synthese in den Schwann-Zellen in Gegenwart von 10% FCS (fötales Kälberserum)
mit und ohne Forskolin zu stimulieren. Einer der getesteten Faktoren
war die GGF-Carboxymethylcellulose-Fraktion
(GGF-CM), die in Gegenwart von FCS, mit und ohne Forskolin mitogenetisch
war. Die Arbeit zeigt, dass in Gegenwart von Forskolin der aus Blutplättchen gewonnene
Wachstumsfaktor (PDGF) ein potentes Mitogen für die Schwann-Zellen ist, wobei
zuvor geglaubt wurde, dass PDGF keinen Effekt auf die Schwann-Zellen
besitzt.
-
Holmes
et al. Science (1992) 256: 1205 und Wen et al. Cell (1992) 69 559
zeigen, dass die DNA-Sequenzen, welche für Proteine codieren, die an
einen Rezeptor (p185erbB2) binden, mit verschiedenen
menschlichen Tumoren assoziiert sind.
-
Das
p185erbB2-Protein ist ein 185 Kilodalton,
die Membran durchdringendes Protein mit Tyrosinkinaseaktivität. Das Protein
wird durch das erbB2 Proto-Onkogen
kodiert (Yarden und Ullrich, Ann. Rev. Biochem. 57: 443(1988)).
Das erbB2-Gen, auch als „HER-2" (in menschlichen
Zellen) und als „neu" (in Rattenzellen)
bezeichnet, ist eng verwandt mit dem Rezeptor für den epidermalen Wachstumsfaktor
(EGF). Kürzliche
Belege zeigen, dass Proteine, die interagieren mit (und die Kinase
aktivieren von) p185erbB2 die Proliferation
in den Zellen, welche p185erbB2 besitzen,
induzieren (Holmes et al., Science 256: 1205 (1992); Dobashi et
al., Proc. Natl. Acad. Sci. 88: 8582 (1991); Lupu et al., Proc.
Natl. Acad. Sci. 89: 2287 (1992)). Es ist ferner offensichtlich,
dass das Gen, welches die p185erbB2 Bindeproteine
codiert, eine Reihe von unterschiedlich großen, unterschiedlich gespleißte RNA-Transkripte
herstellt, die zu einer Serie von Proteinen führen, die unterschiedlich lang
sind und einige gemeinsame Peptidsequenzen und einige einmalige
Peptidsequenzen enthalten. Dies wird unterstützt durch unterschiedlich gespleißte RNA-Transkripte,
die aus menschlichem Brustkrebs gewinnbar sind (MDA-MB-231) (Holmes
et al. Science 256: 1205 (1992)). Eine weitere Unterstützung kommt
von Proteinen mit einem breiten Größenbereich, die (wie hier offenbart)
als Liganden für
den p185erbB2 Rezeptor agieren (siehe unten).
-
Die
Erfindung stellt im Allgemeinen Verfahren für die Stimulierung der Mitogenese
von Gliazellen (insbesondere von Schwann-Zellen und Gliazellen des
Zentralnervensystems) sowie neue Proteine, die solch eine mitogenetische
Aktivität
auf Gliazellen ausüben,
bereit. Ferner wird DNA bereitgestellt, die für diese Proteine und Antikörper codiert,
welche an diese und an verwandte Proteine binden.
-
Die
neuen Proteine gemäß der Erfindung
umfassen alternative Spleißprodukte
von Sequenzen, die für bekannte
Polypeptide codieren. Diese bekannten Proteine sind im Allgemeinen
Mitglieder der GGF/p185erbB2-Proteinfamilie.
-
Die
Erfindung stellt insbesondere Polypeptide mit einer beschriebenen
Formel bereit und DNA-Sequenzen, die für diese Polypeptide codieren.
-
Die
Erfindung stellt ein Polypeptid bereit, das von einem GGF/p185erbB2 Ligandengen codiert wird und mit einem
p185erbB2-Rezeptor interagiert, wobei das
Polypeptid eine E-Domäne,
die von SEQ ID Nr. 163 codiert wird, oder eine E-Domäne, welche
die Aminosäuresequenz
von SEQ ID Nr. 210 umfasst, und eine dem epidermalen Wachstumsfaktor ähnliche
Domäne
umfasst, die
- a) eine C-Nukleinsäuresequenz
(SEQ ID Nr. 177) und entweder eine C/D Nukleinsäuresequenz (SEQ ID Nr. 178)
oder eine C/D' Nukleinsäuresequenz
(SEQ ID Nr. 143) in der 5' zu
3'-Reihenfolge C-C/D
oder C-CD';
- b) die Aminosäuresequenz
von SEQ ID Nr. 151, 152, 220, 221, 222, 223, 224 oder 225; oder
- c) die Aminosäuren
362–411
von SEQ ID Nr. 170
umfasst.
-
Gemäß einem
ersten Aspekt der Erfindung induziert das Polypeptid die Zellteilung
von Gliazellen.
-
Gemäß einem
zweiten Aspekt der Erfindung induziert das Polypeptid die Acetylcholinrezeptor-Synthese
in einer Zelle.
-
Gemäß einem
dritten Aspekt der Erfindung induziert das Polypeptid die Myelinisierung
einer Neuralzelle durch eine Gliazelle.
-
Gemäß einer
bevorzugten Ausführungsform
der Erfindung umfasst das Polypeptid die Aminosäuresequenz von SEQ ID Nr. 170.
-
Die
Erfindung stellt ferner eine isolierte Nukleinsäuresequenz bereit, die für das oben
definierte Polypeptid codiert.
-
Gemäß einer
bevorzugten Ausführungsform
der Erfindung umfasst die Nukleinsäuresequenz SEQ ID Nr. 21.
-
Die
Erfindung umfasst ferner Vektoren, die DNA-Sequenzen umfassen, welche
für die
oben definierten Aminosäuresequenzen
codieren. Ferner umfasst ist eine Wirtszelle, welche die isolierte
DNA enthält,
welche für
die oben definierten Aminosäuresequenzen
codiert. Die Erfindung umfasst ferner solche Verbindungen, die an
den p185erbB2-Rezeptor binden und die Zellteilung
von Gliazellen in vivo und/oder in vitro stimulieren, wie hierin
definiert.
-
Ferner
können
Antikörper
gegen die hier beschriebenen Peptide für die Reinigung der hier beschriebenen
Polypeptide verwendet werden. Die Antikörper gegen die Polypeptide
können
auch für
die therapeutische Hemmung der Zellteilung der Gliazellen verwendet
werden.
-
Die
Erfindung umfasst ferner ein Verfahren für die Herstellung eines mitogenetischen
Faktors für
die Gliazellen, dass die Kultur von modifizierten Wirtszellen, wie
oben definiert, unter Bedingungen umfasst, welche die Expression
der DNA-Sequenzen nach der Erfindung ermöglichen.
-
Ein
Verfahren für
die Stimulierung der Zellteilung von Gliazellen kann bewirkt werden
durch das Inkontaktbringen der Gliazellen mit einem oben definierten
Polypeptid als ein Gliazellenmitogen in vivo oder in vitro. Ein
Verfahren für
das Erzeugen eines mitogenetischen Effektes von Gliazellen in einem
Wirbeltier (vorzugsweise einem Säugetier,
mehr bevorzugt bei Menschen) kann bewirkt werden durch die Verabreichung
einer wirksamen Menge des hier definierten Polypeptides.
-
Ein
Verfahren zur Behandlung oder Prophylaxe für eine Nervenerkrankung oder
Störung
kann mit den hier beschriebenen Polypeptiden erreicht werden. Ein
Verfahren für
die Prophylaxe oder die Behandlung eines pathophysiologischen Zustandes
des Nervensystems, bei dem ein Zelltyp involviert ist, der gegenüber den
hier definierten Polypeptiden empfindlich oder reagierend ist, kann
mit den vorliegenden Polypeptiden auch erzielt werden.
-
Die
erwähnten
Verfahren zur Behandlung umfassen Verfahren, welche die Störung eines
peripheren Nervenschadens beinhaltet, einen Nervenschaden in dem
zentralen Nervensystem, eine neurodegenerative Störung, eine
Demyelinisation im peripheren oder im zentralen Nervensystem oder
ein Schaden oder ein Verlust von Schwann-Zellen, Oligodendrozyte,
Mikroglia oder Astrozyte. Zum Beispiel sind Nervenleiden von sensorischen
oder motorischen Nervenfasern oder die Behandlung einer neurodegenerativen
Störung
mit umfasst. In jedem dieser Fälle
besteht die Behandlung in der Verabreichung einer wirksamen Menge
des Polypeptides.
-
Ein
Verfahren zur Induktion der Nervenregeneration und/oder zur Reparatur
kann auch durch die Verabreichung einer wirksamen Menge des oben
definierten Polypeptides bewirkt werden. Solch ein Medikament wird
hergestellt durch Verabreichung des Polypeptides mit einem pharmazeutisch
wirksamen Träger.
-
Die
Erfindung umfasst die Verwendung eines Polypeptides, wie oben definiert,
bei der Herstellung eines Medikamentes.
-
Die
oben definierten Polypeptide können
verwendet werden
- – um ein Säugetier zu immunisieren für die Produktion
von Antikörpern,
die optional für
therapeutische oder diagnostische Zwecke verwendet werden können,
- – in
einer kompetitiven Prüfung
zur Identifikation oder Quantifizierung von Molekülen mit
Rezeptorbindungseigenschaften, die denen des Polypeptides entsprechen,
und/oder
- – für den Kontakt
einer Probe mit einem Polypeptid, wie oben erwähnt, zusammen mit einem Rezeptor,
der spezifisch an das Polypeptid binden kann, um die kompetitive
Hemmung der Bindung an das Polypeptid nachzuweisen,
- – in
einem Affinitätsisolierungsverfahren,
optional Affinitätschromatographie,
für die
Abtrennung eines entsprechenden Rezeptors.
-
Die
Erfindung umfasst ferner EGFL1, EGFL2, EGFL3, EGFL4, EGFL5 und EGFL6
Polypeptide, die 38 bis 43 und
die SEQ ID Nrn. 220–225
zur Verwendung bei der Stimulierung der Zellteilung von Gliazellen
in vivo und in vitro.
-
Von
der Erfindung ist auch die Verabreichung des GGF-II Polypeptides,
dessen Sequenz in der 45A dargestellt
ist, für
die Stimulierung der Zellteilung von Gliazellen umfasst.
-
Gemäß einem
zusätzlichen
Aspekt der vorliegenden Erfindung können die hier beschriebenen,
neuen Polypeptide dazu verwendet werden, um die Synthese von Acetylcholinrezeptoren
zu stimulieren.
-
Wie
oben erwähnt,
stellt die Erfindung neue Gliawachstumsfaktoren aus Säugetierquellen,
einschließlich
Rind und Mensch, bereit, welche von bekannten Faktoren verschieden
sind. Diese Faktoren sind mitogenetisch für Schwann-Zellen gegenüber einem
Hintergrund von fötalem
Kälberplasma
(FCP). Die Erfindung stellt ferner Verfahren für die Herstellung von diesen
Faktoren bereit sowie ein verbessertes Verfahren für die Definition
der Aktivität
von diesen und anderen Faktoren. Therapeutische Anwendungen der
Faktoren sind ferner ein signifikanter Aspekt der vorliegenden Erfindung.
-
Die
Erfindung umfasst ferner Sequenzen, die eine Sequenzidentität der Homologie
zu den oben angezeigten Sequenzen von mehr als 60%, vorzugsweise
mehr als 80%, haben.
-
Obwohl
die vorliegende Erfindung nicht auf eine besondere Gruppe an Hybridisierungsbedingungen beschränkt ist,
gibt das folgende Protokoll eine allgemeine Anleitung, die befolgt
werden kann, falls dies gewünscht
ist.
-
DNA-Proben
können
mit einer hohen spezifischen Aktivität (etwa 108 bis
109 32Pdmp/μg) durch Nick-Translation
oder durch PCR-Reaktionen gemäß Schowalter
und Sommer (Anal. Biochem., 177: 90–94, 1989) markiert und durch
Entsalzung auf G-150 Sephadex-Säulen
gereinigt werden. Die Proben können
denaturiert (10 Minuten in kochendem Wasser und anschließendem Eintauchen
in Eiswasser) und dann zu Hybridisierungslösungen von 80% Puffer B gegeben
werden (2 g Polyvinylpyrrolidin, 2 g Ficoll-400, 2 g Rinderserumalbumin,
50 ml 1 M Tris HCL (pH 7,5), 58 g NaCl, 1 g Natriumpyrophosphat,
10 g Natriumdodecylsulfat, 950 ml Wasser), der 10% Dextransulfat
enthält,
wobei die Zugabe mit 106 dpm 32P
pro ml erfolgen kann.
-
Es
erfolgt dann eine Inkubation über
Nacht (etwa 16 Stunden) bei 60°C.
Die Filter können
dann bei 60°C
gewaschen werden, zuerst in Puffer B für 15 Minuten und anschließend dreimal
für jeweils
20 Minuten in 2 × SSC,
0,1% SDS und dann einmal für
20 Minuten in 1 × SSC,
0,1% SDS.
-
In
anderer Hinsicht stellt die Erfindung bereit:
- (a)
Einen Grundpolypeptidfaktor, der bei Erzielung aus Rinderhypophysenmaterial
ein beobachtetes Molekulargewicht, ob unter reduzierenden Bedingungen
oder nicht, von etwa 30 kD bis etwa 36 kD auf SDS-Polyacrylamid-Gelelektrophorese
unter Verwendung der folgenden Molekulargewichtsstandards hat:
| Lysozym
(Hühnereiweiß) | 14.400 |
| Sojabohnen-Trypsininhibitor | 21.500 |
| Carboanhydrase
(Rind) | 31.000 |
| Ovalbumin
(Hühnereiweiß) | 45.000 |
| Rinderserumalbumin | 66.200 |
| Phosphorylase
B (Kaninchenmuskel) | 97.400; |
wobei der Faktor eine mitogenetische Aktivität auf Gliazellen
hat, einschließlich
der Stimulierung der Teilung von Schwann-Zellen der Ratte in Gegenwart
von fötalem
Kälberplasma,
und bei Isolierung unter Verwendung der Hochdruck-Flüssigkeitschromatographie
an Umkehrphasen wenigstens 50% dieser Aktivität nach 10 Wochen Inkubation
in 0,1% Trifluoressigsäure
bei 4°C
behält;
und - (b) ein Grundpolypeptidfaktor, der bei Erzielung aus Rinderhypophysenmaterial
ein beobachtetes Molekulargewicht unter nicht-reduzierenden Bedingungen von etwa 55
kD bis etwa 63 kD auf SDS-Polyacrylamid-Gelelektrophorese
unter Verwendung der folgenden Molekulargewichtsstandards zeigt:
| Lysozym
(Hühnereiweiß) | 14.400 |
| Sojabohnen-Trypsininhibitor | 21.500 |
| Carboanhydrase
(Rind) | 31.000 |
| Ovalbumin
(Hühnereiweiß) | 45.000 |
| Rinderserumalbumin | 66.200 |
| Phosphorylase
B (Kaninchenmuskel) | 97.400; |
wobei das menschliche Äquivalent von diesem Faktor
durch den hierin beschriebenen DNA-Klon GGF2HBS5 codiert wird, und
dieser Faktor eine mitogenetische Aktivität auf Gliazellen hat, einschließlich der
Stimulierung der Teilung von Schwann-Zellen der Ratte in Gegenwart
von fötalem
Kälberserum,
und bei Isolierung unter Verwendung der Hochdruck-Flüssigkeitschromatographie
an Umkehrphasen wenigstens 50% der Aktivität nach 4 Tagen Inkubation in
0,1% Trifluoressigsäure
bei 4°C
behält.
-
Nur
zur Darstellung in dieser Beschreibung werden die Niedrigmolekulargewichtsfaktoren
und die Hochmolekulargewichtsfaktoren gemäß der vorliegenden Erfindung
im Folgenden als „GGF-I" und „GGF-II" bezeichnet. Die „GGF2" Bezeichnung wird
für alle
Klone verwendet, die mit den Peptidsequenzdaten isoliert wurden,
welche von dem GGF-II Protein abstammen (das heißt, GGF2HBS5, GGF2BPP3).
-
Es
sei angemerkt, dass die Grenzen der Molekulargewichtsbereiche nicht
exakt anzugeben sind, sondern Gegenstand von leichten Variationen
in Abhängigkeit
von der Quelle des einzelnen Polypeptidfaktors ist. Eine Variation
von etwa 10% würde
zum Beispiel nicht unmöglich
sein für
ein Material aus einer anderen Quelle.
-
Ein
anderer Aspekt der vorliegenden Erfindung benutzt die Tatsache,
dass die Gliawachstumsfaktoren und die p185erbB2 Ligandenproteine
von dem gleichen Gen codiert werden. Eine Reihe von messenger-RNA Spleißvarianten
(und die daraus resultierenden Proteine) stammen von diesem Gen
ab und viele von diesen Produkten zeigen eine p185erbB2 Bindung
und Aktivierung. Verschiedene (GGF-II) Genprodukte zeigen eine mitogenetische
Aktivität
für Schwann-Zellen.
Die Erfindung stellt die Verwendung von all diesen bekannten Produkten
des GGF/p185erbB2 Ligandengens (beschrieben
in den oben aufgeführten
Referenzen) als Schwann-Zellmitogene bereit.
-
Diese
Erfindung bezieht sich auch auf andere, noch nicht natürlich isolierte
Spleißvarianten
des Gens für
den Gliawachstumsfaktor. Die 30 zeigt
das bekannte Muster des Spleißens,
welches von Polymerase-Kettenreaktions-Experimenten
abstammt (auf umgekehrt transkribierter RNA) und die Analyse der
cDNA-Klone (wie hier dargestellt), die abstammen von Sequenzen,
die publiziert sind als Sequenzen, welche p185erbB2 Liganden
codieren (Peles et al., Cell 69: 205 (1992) und Wen et al., Cell
69: 559 (1992)). Diese Muster sowie die zusätzlichen, hier offenbarten
Muster stellen vermutlich Spleißvarianten
dar, die existieren. Ein weiterer Aspekt der vorliegenden Erfindung
bezieht sich auf Nukleotidsequenzen, die neue Proteinfaktoren codieren,
welche von diesem Gen abstammen. Die Erfindung stellt ferner Verfahren
für die
Herstellung von diesen Faktoren bereit. Die therapeutische Anwendung
von diesen neuen Faktoren ist ein weiterer Aspekt der Erfindung.
-
Diese
anderen wichtigen Aspekte der Erfindung sind:
Eine Reihe von
Polypeptidfaktoren, die mitogenetische Aktivität für Gliazellen besitzen, einschließlich der
Stimulierung der Teilung von Schwann-Zellen, und deren Reinigung und Charakterisierung
gemäß den Verfahren,
die dargestellt sind durch Lupu et al., Science 249: 1552 (1990);
Lupu et al., Proc. Natl. Acad. Sci USA, 89: 2287 (1992); Holmes
et al., Science 256: 1205 (1992); Peles et al. 69: 205 (1992); Yarden
und Peles, Biochemistry 30: 3543 (1991); Dobashi et al., Proc. Natl.
Acad. Sci. 88: 8582 (1991); Davin et al., Biochem. Biophys. Res.
Commun. 179: 1536 (1991); Beaumont et al., Patentanmeldung PCT/US
91/03443 (1990); Greene et al., Patentanmeldung PCT/US 91/02331
(1990); Usdin und Fischbach, J. Cell. Biol. 103: 493–507 (1986); Falls
et al., Cold Spring Harbor Symp. Quant. Biol. 55: 397–406 (1990);
Harris et al., Proc. Natl. Acad. Sci. USA 88: 7664–7668 (1991)
und Falls et al., Cell 72: 801–815
(1993).
-
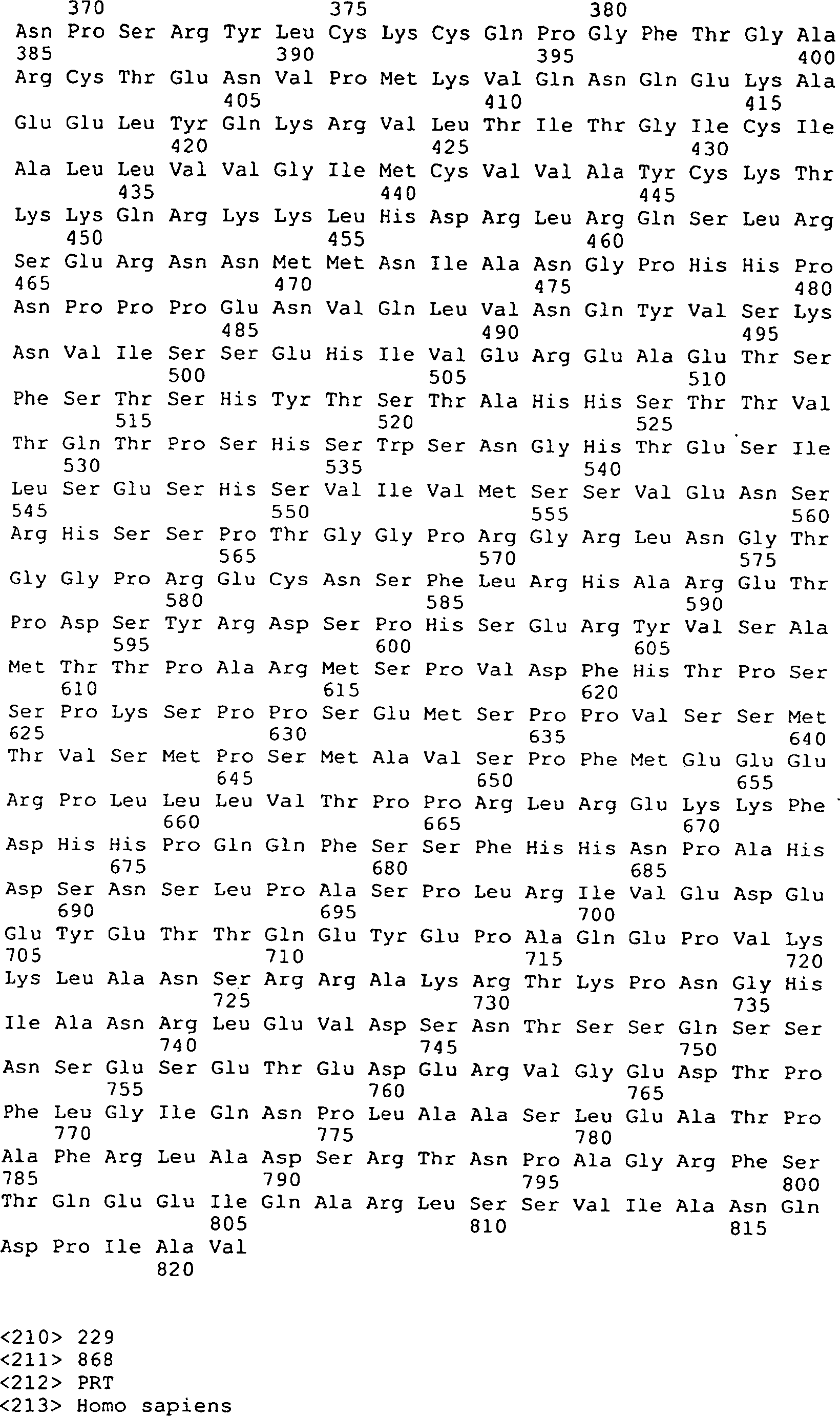
Die
menschlichen Peptidsequenzen, die oben beschrieben und in den 31, 32, 33 und 34 (SEQ
ID Nrn. 192, 194, 195 und 147–219)
dargestellt sind, stellen eine Reihe von Spleißvarianten dar, die aus vollständigen komplementären DNAs
(cDNAs) aus natürlichen
Quellen (cDNA-Bibliotheken, hergestellt mit den geeigneten Geweben)
isoliert werden können
oder die als DNA-Konstrukte mit einzelnen Exons (zum Beispiel abstammend
von separaten Exons) durch Fachleute zusammengesetzt werden können.
-
Andere
Verbindungen, insbesondere Peptide, die spezifisch an den p185erbB2-Rezeptor binden, können auch als Gliazellenmitogen
verwendet werden. Eine Kandidatenverbindung kann routinemäßig für eine p185erbB2 Bindung überprüft werden und bei Bindung kann
sie dann auf mitogenetische Aktivität für Gliazellen unter Verwendung
der hier beschriebenen Verfahren getestet werden.
-
Die
Erfindung umfasst jegliche Modifikationen oder Äquivalente der obigen Polypeptidfaktoren,
sofern sie keine signifikant reduzierte Aktivität zeigen. Modifikationen, bei
denen der Aminosäuregehalt
oder die Aminosäuresequenz
verändert
sind, ohne das im Wesentlichen die Aktivität nachteilig beeinflusst wird,
sind umfasst. Zur Darstellung sei auf die EP-A-109748 verwiesen, wo Mutationen von
nativen Proteinen offenbart sind, bei denen die Möglichkeit
einer nicht gewünschten
Disulfidbindung dadurch vermieden ist, dass Cystein, das für die biologische
Aktivität
nicht notwendig ist, durch eine neutrale Aminosäure in der nativen Sequenz
ersetzt ist. Die Feststellungen zur Wirkung und Verwendung, wie
sie hier getroffen werden, treffen somit auch auf solche Verwendungen
und Wirkungen zu, welche modifizierte oder äquivalente Faktoren, die Teil
der Erfindung sind, verwenden.
-
Die
neuen Sequenzen gemäß der Erfindung
unterliegen den Vorteilen der rekombinanten Technologie. Die Erfindung
umfasst somit auch die folgenden Aspekte:
- (a)
DNA-Konstrukte, welche die oben definierten DNA-Sequenzen umfassen,
in operablen offenen Leserahmpositionen innerhalb von Vektoren (benachbart
angeordnet zu Steuersequenzen, um die Expression der Sequenzen zu
ermöglichen)
in ausgewählten
Wirtszellen nach deren Transformation durch die Konstrukte (vorzugsweise
umfasst die Steuersequenz regulierbare Promotoren, zum Beispiel
Trp). Es sei angemerkt, dass die Auswahl eines Promotors und der
regulatorischen Sequenzen, falls notwendig, Gegenstände sind,
die ein Fachmann auswählen
kann;
- (b) Wirtszellen, die durch die Einlagerung der in (a) definierten
Konstrukte modifiziert sind, so dass diese DNA-Sequenzen in diesen
Wirtszellen exprimiert werden können.
Die Wahl des Wirtes ist nicht kritisch und die ausgewählten Zellen
können
prokaryontisch oder eukaryontisch sein und können genetisch modifiziert sein,
um diese Konstrukte durch bekannte Verfahren einzubauen; und
- (c) Verfahren für
die Herstellung der oben definierten Faktoren, welches die Kultur
der modifizierten Wirtszellen unter Bedingungen umfasst, welche
die Expression der DNA-Sequenzen erlauben. Für jede besondere Ausführungsform
können
diese Bedingungen von Fachleuten auf dem Gebiet der rekombinanten DNA-Technologie
vollständig
bestimmt werden. Gliazellmitogene, die durch diese Mittel hergestellt
werden, sind von der vorliegenden Erfindung umfasst. Keiner der
im Stand der Technik beschriebenen Faktoren hat die Kombination
an Eigenschaften, welche die vorliegenden neuen Polypeptidfaktoren
besitzen.
-
Wie
dargelegt, verwendet der Schwann-Zelltest, der für die vorliegenden Faktoren
zur Charakterisierung verwendet wird, einen Hintergrund aus fötalem Kalbsplasma.
In allen anderen Aspekten kann der Test der gleiche sein, der von
Brockes et al. in Meth. Enz., siehe oben, beschrieben ist, wobei
aber 10% FCS ersetzt ist durch 10% FCP. Dieser Unterschied in der
Testtechnik ist wichtig, da die Abwesenheit der aus Blutplättchen gewonnen Faktoren
im fötalen
Kalbsplasma (wie entgegensetzt zum Serum) eine strengere Definition
der Affinität
auf die Schwann-Zellen ermöglicht,
da mögliche
falsche Wirkungen von einigen anderen Faktoren eliminiert sind.
-
Die
Erfindung umfasst auch ein Verfahren für die Herstellung eines oben
definierten Polypeptides, wobei aus Wirbeltiergehirnmaterial extrahiert
wird, um das Protein zu erzielen. Der erzielte Extrakt wird einer
chromatographischen Reinigung mittels Hydroxyapatit-HPLC unterworfen
und dann wird mit diesen Fraktionen eine SDS-Polyacrylamid-Gelelektrophorese
durchgeführt.
Die Fraktion, die ein beobachtetes Molekulargewicht von etwa 30
kD bis 36 kD hat, und/oder die Fraktion, die ein beobachtetes Molekulargewicht
von etwa 55 kD bis 63 kD hat, werden gesammelt. In jedem Fall wird
die Fraktion einer SDS-Polyacrylamid-Gelelektrophorese unter Verwendung
der folgenden Molekulargewichtsstandards unterworfen:
| Lysozym
(Hühnereiweiß) | 14.400 |
| Sojabohnen-Trypsininhibitor | 21.500 |
| Carboanhydrase
(Rind) | 31.000 |
| Ovalbumin
(Hühnereiweiß) | 45.000 |
| Rinderserumalbumin | 66.200 |
| Phosphorylase
B (Kaninchenmuskel) | 97.400 |
-
Im
Falle der kleineren Molekulargewichtsfraktion läuft das SDS- Polyacrylamidgel unter nicht reduzierenden
Bedingungen, unter reduzierenden Bedingungen, oder im Falle der
größeren Molekulargewichtsfraktion
unter nicht reduzierenden Bedingungen. Die Fraktionen werden dann
hinsichtlich ihrer Aktivität
zur Stimulierung der Teilung von Schwann-Zellen der Ratte gegenüber einem
Hintergrund von fötalem
Kalbsplasma getestet.
-
Vorzugsweise
startet das obige Verfahren mit der Isolierung einer relevanten
Fraktion, die durch Carboxymethylcellulosechromatographie erzielt
wird, das heißt,
Material von der Rinderhypophyse. Es ist auch bevorzugt, das Hydroxylapatit-HPLC,
Kationenaustauscherchromatographie, Gelfiltration und/oder einer Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen vor der SDS-Polyacrylamid-Gelelektrophorese
durchgeführt
wird. Bei jedem Stadium in dem Verfahren kann die Aktivität bestimmt
werden unter Verwendung der Einlagerung von radioaktivem Ioddeoxyuridin
in die Schwann-Zellen
als eine Maßnahme
in dem Test, der allgemein beschrieben ist von Brockes in Meth.
Enz., siehe oben, wobei aber 10% FCS ersetzt ist durch 10% FCP.
Wie bereits angemerkt, ist solch ein Test ein Aspekt der Erfindung
für die
mitogenetischen Wirkungen einer Substanz auf die Zellen des zentralen Nervensystems
oder des peripheren Nervensystems, zum Beispiel Schwann-Zellen.
-
Die
Erfindung umfasst somit auch einen Test für die mitogenetische Aktivität von Gliazellen,
wobei ein Hintergrund aus fötalem
Kalbsplasma verwendet wird, um die DNA-Synthese in Gliazellen zu
testen, die durch eine Testsubstanz (wenn überhaupt) stimuliert sind.
-
Ein
weiterer Aspekt der Erfindung ist eine pharmazeutische Formulierung
oder Veterinärformulierung, die
einen der oben definierten Faktoren umfasst, der für die pharmazeutische
oder Veterinärverwendung
formuliert ist und zwar optional zusammen mit einem akzeptablen
Verdünnungsmittel,
Träger
oder Bindemittel und/oder in einer Einheitsarzneiform. Bei Verwendung
der Faktoren gemäß der vorliegenden
Erfindung kann die konventionelle pharmazeutische oder Veterinärpraxis
angewandt werden, um geeignete Formulierungen oder Zusammensetzungen
bereitzustellen.
-
Die
Formulierungen gemäß der vorliegenden
Erfindung können
somit angewandt werden für
die parenterale Verabreichung, zum Beispiel die intravenöse, subkutane,
intramuskuläre,
intraorbitale, opthalmische, intraventrikuläre, intrakraniale, intrakapsuläre, intraspinale,
intracisternale, intraperitoneale, lokale, intranasale, Aerosol-,
Skarifikation- und auch orale, bukkale, rektale oder vaginale Verabreichung.
-
Die
Formulierungen gemäß der vorliegenden
Erfindung können
auch durch Transplantation in den Patienten von Wirtszellen verabreicht
werden, welche die DNA der vorliegenden Erfindung exprimieren, oder durch
Verwendung von chirurgischen Implantaten, welche die Formulierungen
der Erfindung freisetzen.
-
Parenterale
Formulierungen können
in der Form von Flüssiglösungen oder
Suspensionen vorliegen. Für
die orale Verabreichung können
die Formulierungen in der Form von Tabletten oder Kapseln vorliegen.
Für die
intranasale Formulierung können
Pulver, Nasentropfen oder Aerosole benutzt werden.
-
Im
Stand der Technik gut bekannte Verfahren für die Herstellung von Formulierungen
sind zum Beispiel dargestellt in „Remington's Pharmaceutical Sciences". Formulierungen
für die
parenterale Verabreichung können
zum Beispiel als Bindemittel steriles Wasser oder Salzlösung enthalten.
Polyalkylenglycole, wie Polyethylenglycol, Öle von pflanzlichem Ursprung
oder hydrierte Naphthalene, biokompatible, bioabbaubare Lactidpolymere
oder Polyoxyethylen-Polyoxypropylen-Copolymere können verwendet werden, um die
Freisetzung der vorliegenden Faktoren zu steuern. Andere potentiell nützliche,
parenterale Abgabesysteme für
die Faktoren umfassen Ethylen-Vinylacetat-Copolymer-Partikel,
osmotische Pumpen, implantierbare Infusionssysteme und Liposomen.
Formulierungen für
die Inhalation können
zum Beispiel als Bindemittel Laktose enthalten und können wässrige Lösungen sein,
die zum Beispiel Polyoxyethylen-2-Laurylether, Glycocholat und Deoxycholat
enthalten, oder können ölige Lösungen für die Verabreichung
in Form von Nasentropfen sein oder können intranasal als ein Gel
angewandt werden. Formulierungen für die parenterale Verabreichung
können auch
Glycocholat für
die bukkale Verabreichung, Methoxysalicylat für die rektale Verabreichung
oder Zitronensäure
für die
vaginale Verabreichung enthalten.
-
Die
vorliegenden Faktoren können
als alleiniges aktives Mittel oder können in Kombination mit anderen
aktiven Mitteln verwendet werden, zum Beispiel mit anderen Wachstumsfaktoren,
welche das neuronale Überleben
bei neurologischen Krankheiten erleichtern, oder Peptidase- oder
Proteaseinhibitoren.
-
Die
Konzentration der vorliegenden Faktoren in den Formulierungen gemäß der Erfindung
wird in Abhängigkeit
von einer Reihe von Faktoren variieren, einschließlich der
zu verabreichenden Dosierung und der Art der Verabreichung.
-
Im
Allgemeinen können
die Faktoren der vorliegenden Erfindung in einer wässrigen,
physiologischen Pufferlösung
bereitgestellt werden, die etwa 0,1 bis 10% (Gewicht/Volumen) Verbindung
für die
parenterale Verabreichung enthält.
Allgemeine Dosisbereiche erstrecken sich von etwa 1 mg/kg bis etwa
1 g/kg Körpergewicht
pro Tag. Ein bevorzugter Dosisbereich liegt bei etwa 0,01 mg/kg
bis 100 mg/kg Körpergewicht
pro Tag. Die bevorzugte, zu verabreichende Dosierung wird wahrscheinlich
vom Typ und dem Ausmaß des
Fortschreitens der pathophysiologischen Störung, die zu behandeln ist,
abhängig
sein sowie von dem allgemeinen Gesundheitszustand des Patienten,
der Herstellungsart der Formulierung und der Art der Verabreichung.
-
Wie
oben angezeigt sind die Schwann-Zellen (die Gliazellen des peripheren
Nervensystems) stimuliert, um in Gegenwart der Faktoren gemäß der vorliegenden
Erfindung sich zu teilen. Schwann-Zellen des peripheren Nervensystems
sind beteiligt bei der Unterstützung
der Neuronen und bei der Schaffung der Myelinscheiden um die einzelnen
Nervenfasern. Diese Scheiden sind wichtig für die geeignete Weiterleitung
von elektrischen Impulsen zu den Muskeln und von den sensorischen
Rezeptoren.
-
Es
gibt eine Reihe von peripheren Nervenleiden, bei denen die Schwann-Zellen und die Nervenfasern beschädigt sind
und zwar entweder primär
oder sekundär.
Es gibt viele Nervenleiden von sowohl sensorischen als auch motorischen
Fasern (Adams und Victor, „Principles
of Neurology").
Die wichtigsten dieser Nervenleiden sind wahrscheinlich die Nervenleiden,
die mit Diabetes, multiple Sklerose, Landry-Guillain-Barr-Syndrom assoziiert
sind, Nervenleiden, die durch Karzinome hervorgerufen werden sowie
Nervenleiden, die durch toxische Mittel (einige von ihnen werden
verwendet zur Behandlung von Karzinomen) hervorgerufen werden.
-
Die
Erfindung zielt jedoch allgemein auf die Behandlung oder Prophylaxe
von Zuständen,
bei denen ein Schaden des Nervensystems durch jegliche Ursache entstanden
ist, zum Beispiel durch Infektion oder Verletzung. Zusätzlich zu
der Verwendung der vorliegenden Faktoren bei der Behandlung von
Störungen
oder Krankheiten des Nervensystems, wo eine Demyelinisation oder
ein Verlust von Schwann-Zellen vorliegt, können solche Gliawachstumsfaktoren
auch bei der Behandlung von Störungen
des Nervensystems wertvoll sein, die hervorgerufen werden durch
einen Schaden an den peripheren Nerven. Nach Schädigung von peripheren Nerven
führt der
Regenerationsprozess zu einem Wachstum oder Wiederherstellung der
Schwann-Zellen, wobei sich die Beförderung der Nervenfasern zurück zu ihrem
Ziel anschließt.
Durch Erhöhung
der Teilung der Schwann-Zellen könnte
dieser regenerative Prozess nach einem Schaden gefördert werden.
-
Ähnliche
Ansätze
könnten
benutzt werden, um Verletzungen oder neurodegenerative Erkrankungen des
zentralen Nervensystems (Gehirn und Rückenmark) zu behandeln.
-
Ferner
gibt es eine Reihe von Tumoren der Gliazellen, wobei der häufigste
Tumor wahrscheinlich die Neurofibromatose ist, die einen unregelmäßigen, kleinen
Tumor darstellt, der durch eine Überwachsung
der Gliazellen erzeugt wird. Es wurde ferner gefunden, dass eine
Aktivität,
die GGF sehr ähnlich
ist, in einigen Tumoren der Schwann-Zellen auftritt, wodurch Hemmstoffe
der Wirkung der vorliegenden Faktoren auf ihre Rezeptoren eine Therapie
für einen
Gliatumor darstellen, was die Verabreichung einer wirksamen Menge
einer Substanz umfasst, welche die Bindung eines Faktors, wie oben
definiert, an einen Rezeptor hemmt.
-
Im
Allgemeinen erlaubt die Erfindung die Verwendung der vorliegenden
Polypeptidfaktoren bei der Prophylaxe oder der Behandlung von jeglichem
pathophysiologischen Zustand des Nervensystems, bei dem ein Faktor
sensitiver oder ein auf den Faktor reagierender Zelltyp beteiligt
ist.
-
Die
Polypeptidfaktoren gemäß der vorliegenden
Erfindung können
auch als Immunogene zur Herstellung von Antikörpern, wie monoklonale Antikörper, unter
Benutzung von Standardtechniken verwendet werden. Solche Antikörper werden
von der vorliegenden Erfindung umfasst. Diese Antikörper können andererseits für therapeutische
oder diagnostische Zwecke verwendet werden. Zustände, die wahrscheinlich mit
anormalen Mengen des Faktors assoziiert sind, können durch Verwendung von solchen
Antikörpern
erfasst werden. In vitro Techniken können benutzt werden, wobei
unter Verwendung von Standardverfahren Anwendungstests auf isolierte
Proben fallen. Bildgebende Verfahren, bei denen die Antikörper zum
Beispiel mit radioaktiven Isotopen markiert werden, die außerhalb
des Körpers
dargestellt werden können,
können
unter Verwendung von Techniken hinsichtlich der Art der Tumordarstellung
auch verwendet werden.
-
Die
Erfindung zielt ferner auf die allgemeine Verwendung der vorliegenden
Faktoren als Gliazell-Mitogene in vivo oder in vitro sowie auf Faktoren
für solch
eine Verwendung. Eine spezifische Ausführungsform umfasst die Verabreichung
einer wirksamen Menge eines Faktors gemäß der vorliegenden Erfindung,
insbesondere ein Verfahren für
die Herstellung eines mitogenetischen Gliazelleffektes in einem
Wirbeltier für
die Behandlung oder die Prophylaxe einer Erkrankung oder Störung des
Nervensystems.
-
Ein
weiterer allgemeiner Aspekt der Erfindung besteht in der Verwendung
eines Faktors gemäß der vorliegenden
Erfindung bei der Herstellung eines Medikamentes, vorzugsweise für die Behandlung
einer Nervenerkrankung oder Störung
oder für
die neurale Regeneration oder Reparatur.
-
Von
der Erfindung ist ferner die Verwendung der Faktoren gemäß der vorliegenden
Erfindung in kompetitiven Tests umfasst, um Moleküle mit Rezeptorbindungseigenschaften,
die denen der Polypeptide entsprechen, zu identifizieren oder zu
quantifizieren. Die Polypeptide können markiert werden, optional
mit einem Radioisotop. Ein kompetitiver Test kann sowohl Antagonisten
als auch Agonisten des relevanten Rezeptors identifizieren.
-
Gemäß einem
anderen Aspekt stellt die Erfindung die Verwendung von jedem einzelnen
der Faktoren gemäß der vorliegenden
Erfindung in einem Affinitätsisolierungsverfahren,
optional Affinitätschromatographie, für die Trennung
der entsprechenden Rezeptoren bereit. Solche Verfahren für die Isolierung
von Rezeptoren, die bestimmten Proteinen entsprechen, sind im Stand
der Technik bekannt und eine Reihe von Techniken sind verfügbar und
können
auf die Faktoren gemäß der vorliegenden
Erfindung angewandt werden. Der Leser wird zum Beispiel in Bezug
auf IL-6 und IFNγ verwiesen auf Novick,
D. et al., J. Chromatogr. (1990) 510: 331–7. In Bezug auf das Gonadotropin-Freisetzungshormon
wird verwiesen auf Hazum, E., J. (1990) Chromatogr. 510: 233–8. In Bezug
auf G-CSF-Referenz wird verwiesen auf Fukunaga, R., et al., J. Biol.
Chem., 265: 13386–90. In
Bezug auf den IL-2 Verweis wird verwiesen auf Smart, J. E., et al.,
(1990) J. Invest. Dermatol., 94: 158S–163S, und in Bezug auf den
Verweis auf menschliches IFN-Gamma wird verwiesen auf Stefanos,
S. et al., (1989) J. Interferon Res., 9: 719–30.
-
Die
Zeichnungen werden als erstes beschrieben.
-
Zeichnungen
-
Die 1 bis 8 beziehen
sich auf das Beispiel 1 und werden im folgenden kurz beschrieben:
-
1 ist
das Profil für
ein Produkt von der Carboxymethylcellulose-Chromatographie.
-
2 ist
das Profil für
ein Produkt von der Hydroxylapatit-HPLC.
-
3 ist
das Profil für
ein Produkt von Mono-S-FPLC.
-
4 ist
das Profil für
ein Produkt von der Gelfiltration FPLC.
-
Die 5 und 6 zeigen
die Profile für
zwei, teilweise gereinigte Polypeptidprodukte von der Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen.
-
Die 7 und 8 zeigen
Dosis-Effekt-Kurven für
die GGF-I und GGF-II Fraktionen von HPLC mit umgekehrten Phasen
unter Verwendung von fötalem
Kalbsserum oder einem fötalen
Kalbsplasma-Hintergrund.
-
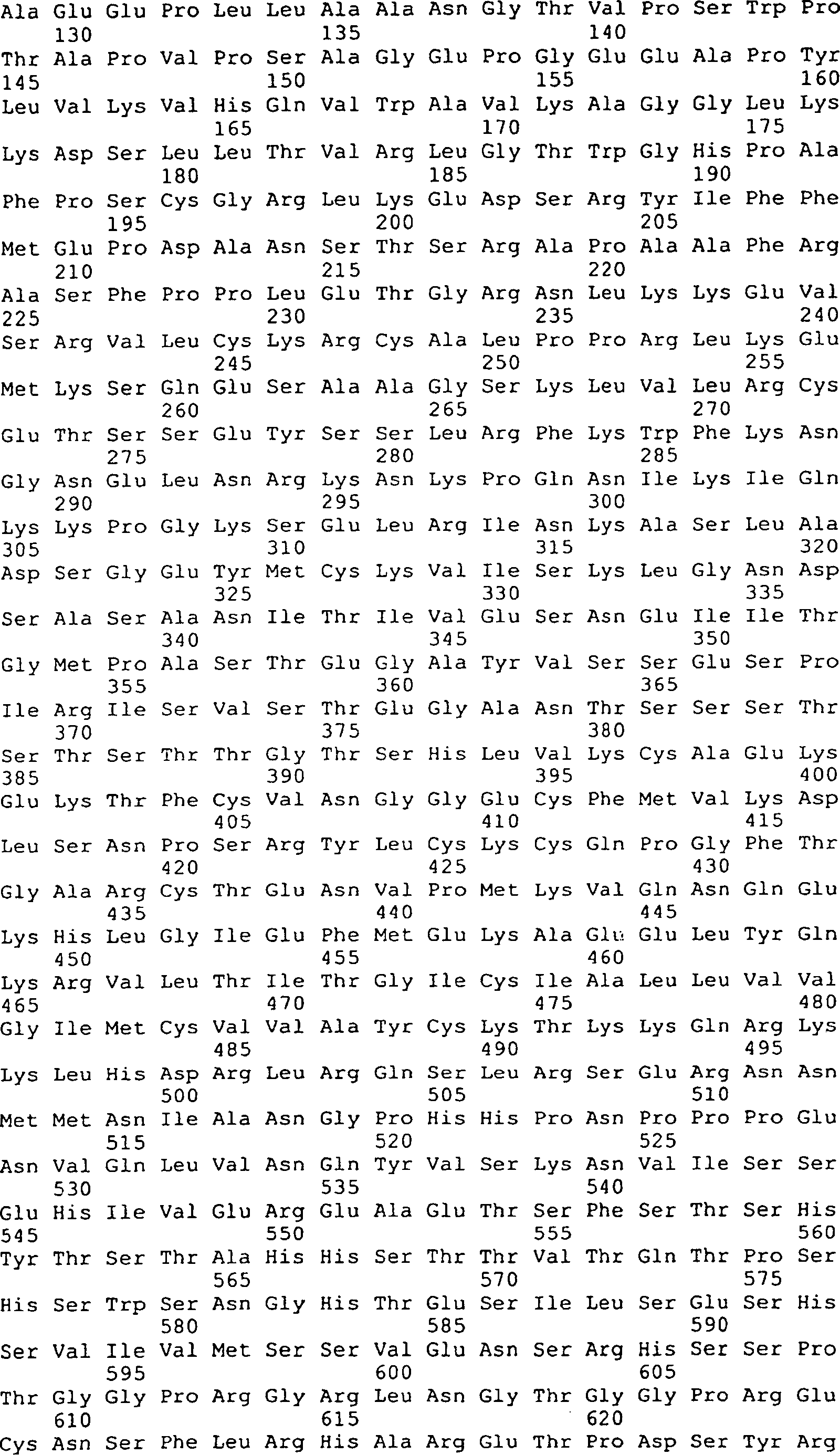
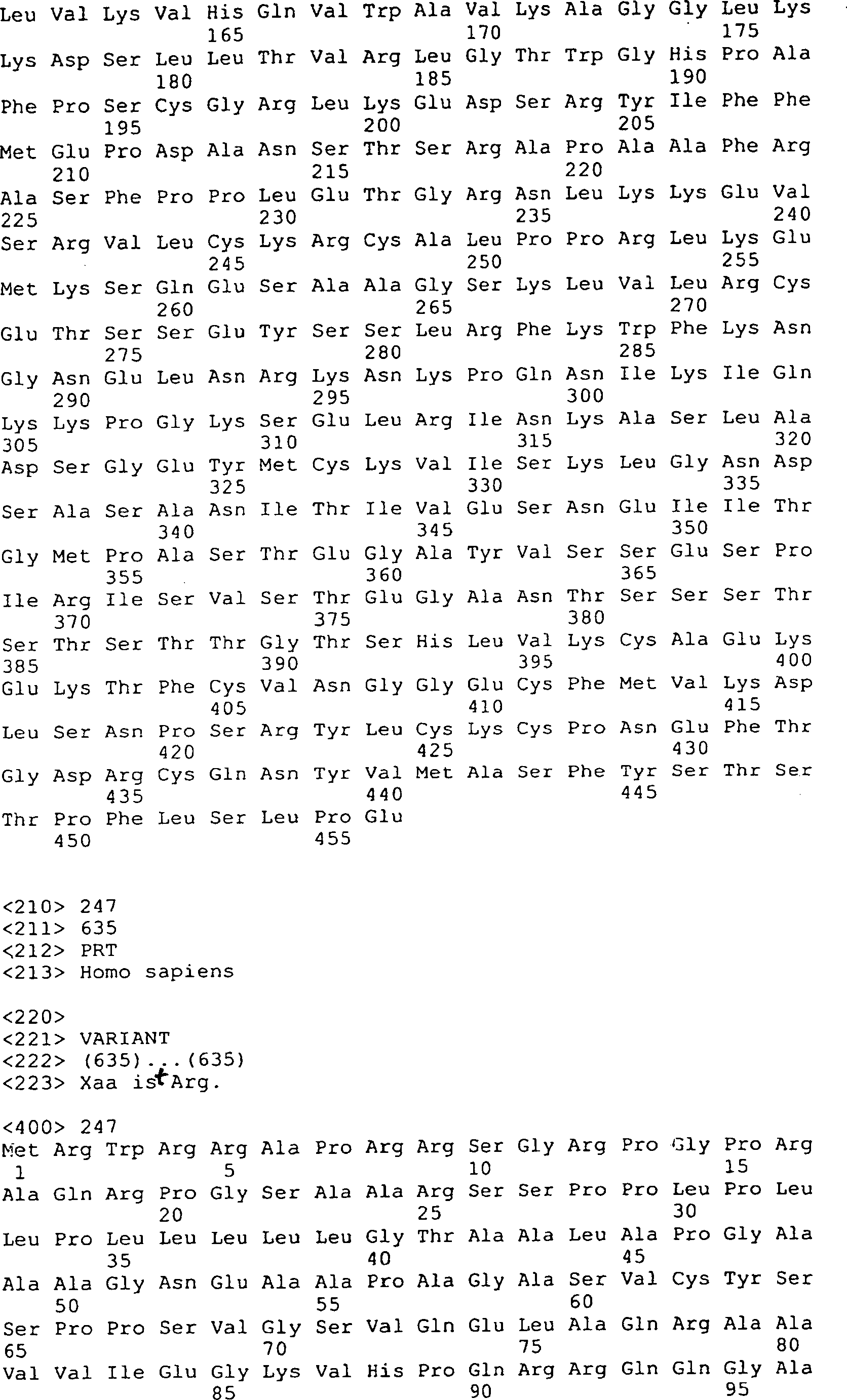
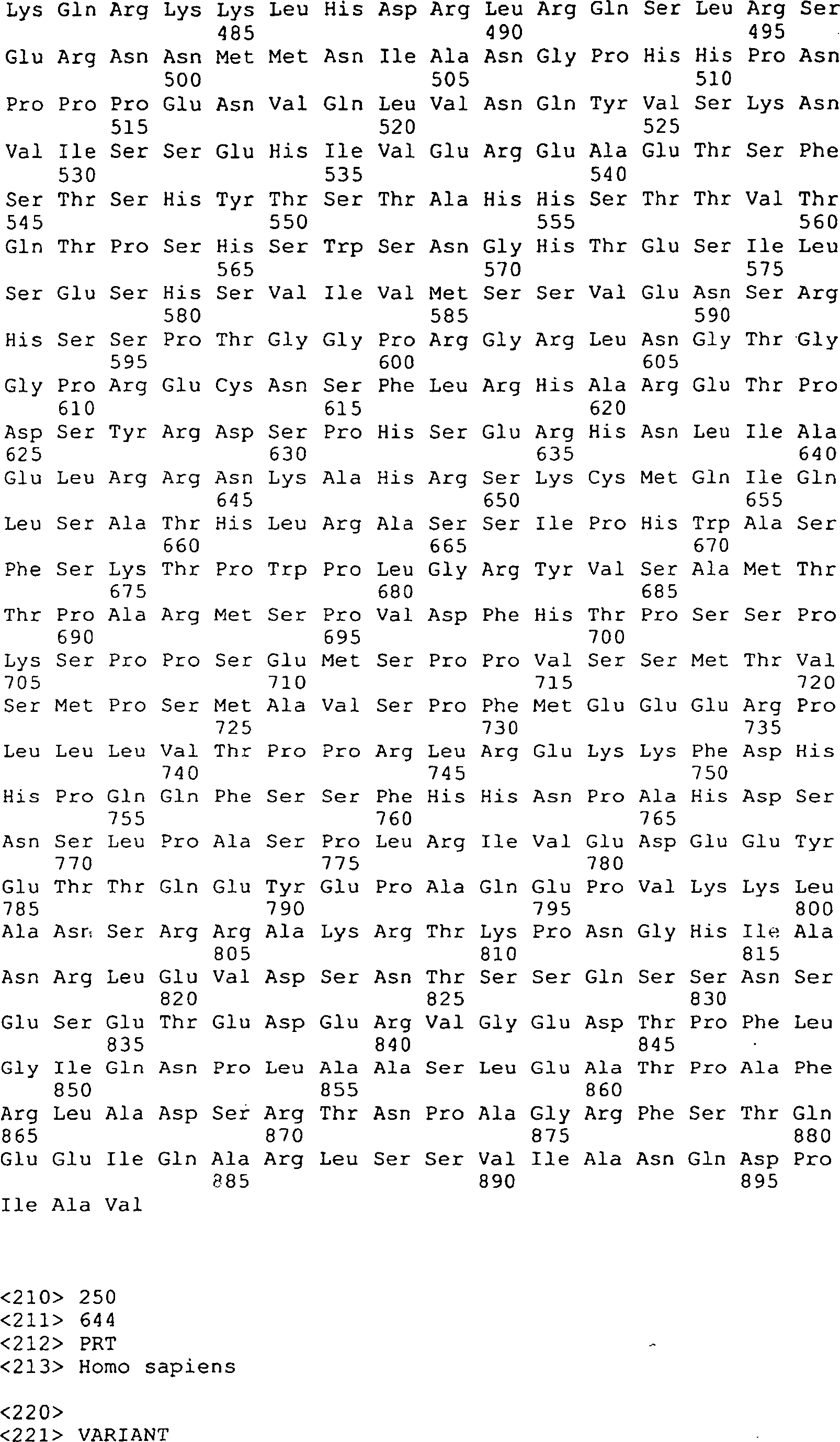
Die 9 bis 12 zeigen
die Peptidsequenzen, die abstammen von GGF-I und GGF-II, SEQ ID Nrn. 1–20, 22–29, 32–41, 43
und 44–169
(siehe folgendes Beispiel 2).
-
9 zeigt
die 21 Peptidsequenzen (SEQ ID Nrn. 1–20 und 169), erzielt von Lysylendopeptidase-
und Protease V8-Verdau von gereinigter Rinderhypophyse GGF-1.
-
In
den 10A und 10B sind
die Sequenzen der GGF-I Peptide aufgeführt, die für die Herstellung der degenerierten
Oligonukleotidproben und degenerierten PCR-Primer verwendet wurden
(SEQ ID Nrn. 1, 22–29 und
17). Einige von den Sequenzen in der 10A wurden
auch für
die Herstellung von synthetischen Peptiden verwendet. Die 10B zeigt die Sequenzen der Peptide, die zu kurz
waren (weniger als 9 Aminosäuren) für die Erstellung
der degenerierten Proben oder degenerierten PCR-Primer (SEQ ID Nrn.
19 und 32).
-
11 zeigt verschiedene Trypsinpeptidsequenzen und
Lysylendopeptidase-C Peptidsequenzen, die abstammen von GGF-II,
SEQ ID Nrn. 33–39,
164–166,
51 und 52.
-
In
der 12 sind unter A die Sequenzen
der GGF-II Peptide gezeigt, die verwendet wurden, um die degenerierten
Oligonukleotidproben und die degenerierten PCR-Primer zu entwerfen
(SEQ ID Nrn. 45–52).
Einige der Sequenzen unter A wurden verwendet, um synthetische Peptide
zu entwerfen. Unter B ist das Peptid gezeigt, das zu kurz war (weniger
als 6 Aminosäuren),
um degenerierte Proben oder degenerierte PCR-Primer (SEQ ID Nr.
53) zu entwerfen.
-
Die 13 bis 20 beziehen
sich auf das Beispiel 3 und veranschaulichen die mitogenetische
Aktivität der
Faktoren gemäß der Erfindung.
-
13 ist ein Diagramm und zeigt den Vergleich der
BrUdR ELISA und der [125I]UdR Zählverfahren für die DNA-Syntheseanalyse
in Schwann-Zellkulturen.
-
Die 14A und 14B sind
Diagramme und zeigen die Br-UdR Immunoreaktivität mit der Zahl der Br-UdR markierten
Zellen.
-
15 zeigt die mitogenetische Antwort von Schwann-Zellen
des Ischiasnervs der Ratte auf GGFs.
-
16 ist ein Diagramm und zeigt die DNA-Synthese
in Schwann-Zellen des Ischiasnervs der Ratte und in 3T3-Fibroplasten
in Gegenwart von GGFs.
-
17 zeigt im Diagramm die mitogenetische Antwort
von BHK 21 C13 Zellen auf FCS und GGFs.
-
18 zeigt im Diagramm das Überleben und die Vermehrung
von BH 21 C13 Zellmikrokulturen nach 48 Stunden in Gegenwart von
GGFs.
-
19 zeigt im Diagramm die mitogenetische Antwort
von C6 Zellen auf FCS.
-
Die 20A und 20B zeigen
im Diagramm die mitogenetische Antwort von C6 Zellen auf aFGF und
GGFs.
-
Die 21 bis 28(a, b und
c) beziehen sich auf das folgende Beispiel 4 und werden im Folgenden
kurz dargestellt:
-
21 ist eine Liste von degenerierten Oligonukleotidproben
(SEQ ID Nrn. 54–76,
78–88),
entworfen von den Peptidsequenzen in der 10 unter
A und der 12 unter A.
-
22 zeigt die mutmaßliche GGF-II Gensequenz vom
Rind von dem rekombinanten genomischen Phagen GGF2BG1 (Rind), enthaltend
die Bindungsstelle der degenerierten Oligonukleotidproben 609 und
650 (siehe die 21, SEQ ID Nrn. 69 und bzw.
72). Die Figur stellt den codierenden Strang der DNA-Sequenz (SEQ
ID Nr. 89) und die abgeleitete Aminosäuresequenz (SEQ ID Nr. 169)
im dritten Leserahmen dar. Die Sequenz von Peptid 12 vom Faktor 2
(Fett) ist Teil eines offenen Leserahmens von 66 Aminosäuren (Nukleotide 75272).
-
23 zeigt die degenerierten PCR-Primer
(SEQ ID Nrn. 90–108)
und die 23B zeigt die PCR-Primer (SEQ
ID Nrn. 109–119),
die in Experimenten verwendet wurden, um Segmente der GGF-II codierenden Sequenzen
(Rind) zu isolieren, die in RNA von dem Hypophysenhinterlappen vorhanden
sind.
-
24 zeigt die neun distinkten, benachbarten GGF-II
cDNA-Strukturen und Sequenzen (Rind), die in PCR-Amplifikationsexperimenten
erzielt wurden unter Verwendung der Primer der 12 unter A und B, und RNA von dem Hypophysenhinterlappen.
Die obere Linie der Figur zeigt schematisch die codierenden Sequenzen,
welche zu den cDNA-Strukturen beitragen, die charakterisiert wurden.
-
25 ist eine physikalische Karte des rekombinanten
Phagen von GGF2BG1 (Rind). Das Rindfragment hat eine Länge von
etwa 20 kb und enthält
zwei Exons (Fett) des GGF-II Gens vom Rind. Die Restriktionsstellen
für die
Enzyme Xbal, SpeI, Ndel, EcoRI, Kpnl und SstI sind auf dieser physikalischen
Karte angegeben. Die schraffierten Abschnitte entsprechen den Fragmenten,
die für
die Sequenzierung subkloniert wurden.
-
26 zeigt die schematische Struktur von drei alternativen
Genprodukten des mutmaßlichen
GGF-II Gens vom Rind. Die Exons sind von A bis E in der Reihenfolge
ihrer Entdeckung gekennzeichnet. Die alternativen Spleißmuster
1, 2 und 3 erzeugen drei überlappende,
abgeleitete Proteinstrukturen (GGF2BPP1, 2 und 3), die in den verschiedenen 28a b, c (unten beschrieben) angezeigt sind.
-
27 (SEQ ID Nrn. 120–132, 45, 52 und 53) ist ein
Vergleich der GGF-I und der GGF-II Sequenzen, identifiziert in den
abgeleiteten Proteinsequenzen, die in den 28A–28E gezeigt sind (siehe unten), mit den Peptidsequenzen,
die in den 10 und 12 aufgeführt sind.
Die Figur zeigt, dass sechs von neun GGF-II Peptidsequenzen in diesen
abgeleiteten Proteinsequenzen berücksichtigt sind. Zwei Peptidsequenzen,
die ähnlich
zu den GGF-I Sequenzen sind, wurden auch gefunden.
-
28A zeigt die DNA-Sequenz des codierenden Stranges
(SEQ ID Nr. 133) und die abgeleitete Aminosäuresequenz (SEQ ID Nr. 190)
der cDNA, erzielt von dem Spleißmuster
Nr. 1 in der 26. Diese partielle cDNA des
mutmaßlichen
Rind GGF-II Gens codiert ein Protein von 206 Aminosäuren. Peptide
in Fettdruck waren solche, die von den Listen der 10 und 12 identifiziert
wurden. Potentielle Glykosilierungsstellen sind unterstrichen (zusammen
mit dem Polyadenylierungssignal AATAAA) (SEQ ID Nr. 189).
-
Die 28B und 28C zeigen
die DNA-Sequenz des codierenden Stranges (SEQ ID Nr. 134) und die
abgeleitete Aminosäuresequenz
(SEQ ID Nr. 191) der cDNA, erzielt aus dem Spleißmuster Nummer 2 der 26. Diese partielle cDNA des mutmaßlichen
Rind GGF-II Gens codiert für
ein Protein von 281 Aminosäuren.
Peptide im Fettdruck sind solche, die von den Listen der 10 und 12 identifiziert
wurden. Potentielle Glykosilierungsstellen sind unterstrichen (zusammen
mit dem Polyadenylierungssignal AATAAA) (SEQ ID Nr. 189).
-
Die 28D und 28E zeigen
die DNA-Sequenz des codierenden Stranges (SEQ ID Nr. 135) und die
abgeleitete Aminosäuresequenz
(SEQ ID Nr. 193) der cDNA, erzielt aus dem Spleißmuster Nummer 3 in der 26. Diese partielle cDNA des mutmaßlichen
Rind GGF-II Gens codiert für
ein Protein mit 257 Aminosäuren.
Peptide im Fettdruck sind solche, die von den Listen der 10 und 12 identifiziert
wurden. Potentielle Glykosilierungsstellen sind unterstrichen (zusammen
mit dem Polyadenylierungssignal AATAAA) (SEQ ID Nr. 189).
-
29, die sich auf das folgende Beispiel 6 bezieht,
ist ein Autoradiogramm einer Kreuzhybridisierungsanalyse der mutmaßlichen
GGF-II Rindergensequenzen mit einer Reihe von Säugetier DNAs auf einem Southern
Blot. Der Filter enthält
Linien von mit EcoRI geschnittener DNA (5 μg pro Reihe) von den in der
Figur aufgeführten
Arten. Die Probe detektiert ein einzelnes starkes Band in jeder
DNA-Probe, einschließlich
eines vier Kilobasen Fragmentes in der Rind-DNA, wie dies durch
die physikalische Mappe von 25 erwartet
wurde. Banden mit relativ geringer Intensität wurden auch beobachtet und
diese könnten
verwandte DNA-Sequenzen darstellen. Das starke Hybridisierungsband
von jeder der anderen Säugetier-DNA-Proben
stellt vermutlich das GGF-II Homologe von diesen Arten dar.
-
30 ist ein Diagramm von beispielhaften Spleißvarianten.
Die codierenden Segmente sind dargestellt durch F, E, B, A, G, C,
C/D, C/D', D, D', H, K und L. Die
Lokalisierung der Peptidsequenzen, die von dem gereinigten Protein
abstammen, sind angezeigt durch „o".
-
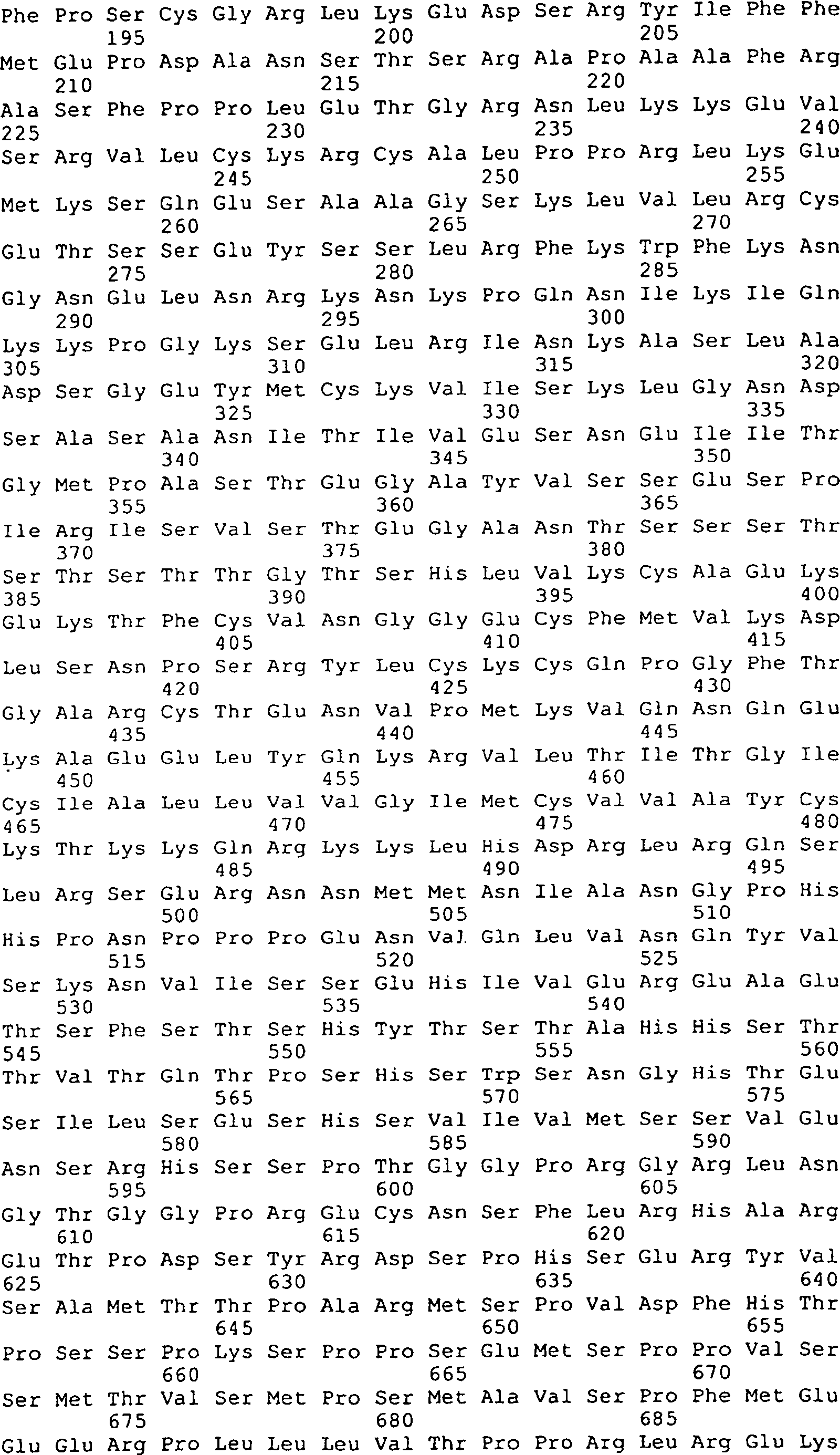
Die 31A–R
zeigen DNA-Sequenzen (SEQ ID Nrn. 42, 44, 77, 136–147, 160,
161, 163 und 173–210)
und die vorhergesagten Peptidsequenzen der Codierungssegmente von
GGF. Die Linie 1 zeigt die vorhergesagten Aminosäuresequenzen von Rind-GGF,
die Linie 2 zeigt die Nukleotidsequenzen von Rind-GGF, die Linie
3 zeigt die Nukleotidsequenzen von menschlichem GGF (Heregulin)
(Nukleotidbasenübereinstimmungen
sind mit einer vertikalen Linie angezeigt), und Linie 4 zeigt die
vorhergesagten Aminosäuresequenzen
von menschlichen GGF/Heregulin, wo es von der vorhergesagten Rindsequenz
abweicht. Die codierenden Segmente E, A' und K stellen nur die Rindsequenzen
dar. Das codierende Segment D' stellt
nur die menschliche (Heregulin) Sequenz dar.
-
Die 32A–B
zeigen die vorhergesagte GGF2 Aminosäuresequenz und die Nukleotidsequenz
von BPP5 (SEQ ID Nr. 195 und SEQ ID Nr. 148). Die obere Linie ist
die Nukleotidsequenz und die untere Linie ist die vorhergesagte
Aminosäuresequenz.
-
Die 33A–B
zeigen die vorhergesagte Aminosäuresequenz
und die Nukleotidsequenz von GGF2BPP2 (SEQ ID Nr. 149 und SEQ ID
Nr. 142). Die obere Linie ist die Nukleotidsequenz und die untere Linie
ist die vorhergesagte Aminosäuresequenz.
-
Die 34A–C
zeigen die vorhergesagte Aminosäuresequenz
und die Nukleotidsequenz von GGF2BPP4 (SEQ ID Nr. 194 und SEQ ID
Nr. 150). Die obere Linie ist die Nukleotidsequenz und die untere Linie
ist die vorhergesagte Aminosäuresequenz.
-
35 (SEQ ID Nrn. 151–152) zeigt die Ausrichtung
der zwei GGF Peptidsequenzen (GGF2bpp4 und GGF2bpp5) mit der menschlichen
EGF (hEGF). Sternchen zeigen die Stellen der konservierten Cysteine
an.
-
36 zeigt die Höhe
der GGF-Aktivität
(mitogenetischer Test mit Schwann-Zellen) und der Tyrosinphosphorilierung
eines etwa 200 kD Proteins (Intensität einer 200 kD Bande auf einem
Autoradiogramm eines Western Blots, entwickelt mit einem polyklonalen
Antiphosphotyrosin-Antikörper)
in Reaktion auf zunehmende Mengen an GGF.
-
Die 37A–B
zeigen die Spleißvariante,
die von den in der 31A–R gezeigten Sequenzen abstammen.
-
38 zeigt die vorhergesagte Aminosäuresequenz
(unten; SEQ ID Nr. 220) und die Nukleinsäuresequenz (oben; SEQ ID Nr.
154) von EGFL1.
-
39 zeigt die vorhergesagte Aminosäuresequenz
(unten; SEQ ID Nr. 221) und die Nukleinsäuresequenz (oben; SEQ ID Nr.
155) von EGFL2.
-
40 zeigt die vorhergesagte Aminosäuresequenz
(unten; SEQ ID Nr. 222) und die Nukleinsäuresequenz (oben; SEQ ID Nr.
156) von EGFL3.
-
41 zeigt die vorhergesagte Aminosäuresequenz
(unten; SEQ ID Nr. 223) und die Nukleinsäuresequenz (oben; SEQ ID Nr.
157) von EGFL4.
-
42 zeigt die vorhergesagte Aminosäuresequenz
(unten; SEQ ID Nr. 224) und die Nukleinsäuresequenz (oben; SEQ ID Nr.
158) von EGFL5.
-
43 zeigt die vorhergesagte Aminosäuresequenz
(unten; SEQ ID Nr. 225) und die Nukleinsäuresequenz (oben; SEQ ID Nr.
159) von EGFL6.
-
44 ist eine maßstabgerechte Mappe des codierenden
Segmentes des Klons. T3 bezieht sich auf den verwendeten Bakteriophagen-Promotor
zur Herstellung von mRNA von dem Klon. R = flankierende Stelle für das Restiktionsenzym
EcoRI. 5' UT bezieht
sich auf die 5' nicht-translatierte
Region. E, B, A, C, C/D' und
D beziehen sich auf die codierenden Segmente. 0 = Start der Translation.
A = 5' Grenze der
Region, die zu dem Rinder E-Segment
homolog ist (siehe Beispiel 6) und 3' UT bezieht sich auf die 3' nicht-translatierte Region.
-
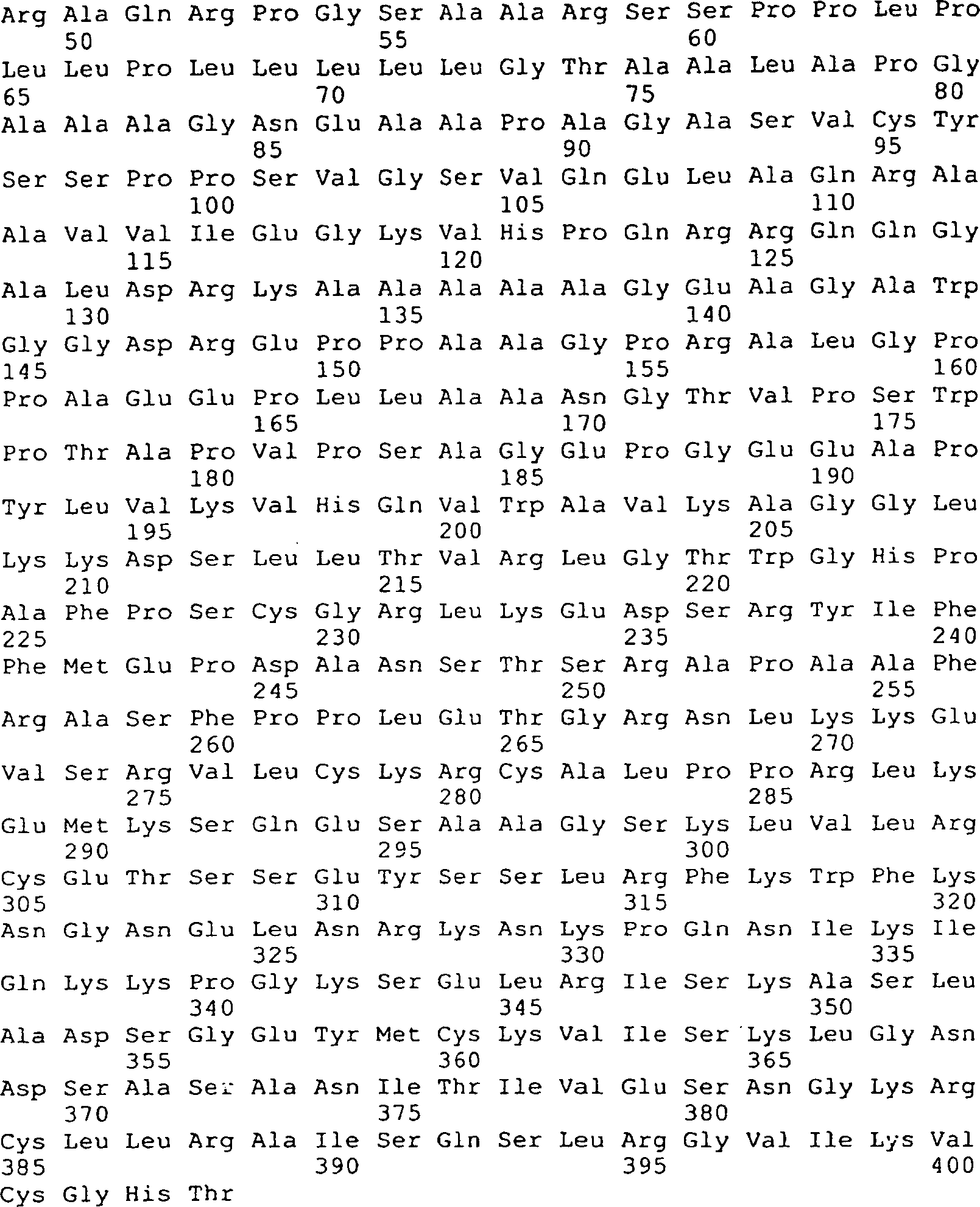
Die 45A–D
zeigen die vorhergesagte Aminosäuresequenz
(in der Mitte; SEQ ID Nr. 170) und die Nukleinsäuresequenz (oben; SEQ ID Nr.
21) von GGF2HBS5. Die untere (mit Unterbrechung) Sequenz stellt die
Peptidsequenzen dar, die von den GGF-II Zubereitungen abstammen
(siehe die 11, 12).
-
46 ist eine graphische Darstellung der mitogenetischen
Aktivität
von rekombinanten, menschlichen und vom Rind abstammenden Gliawachstumsfaktoren
auf Schwann-Zellen.
-
47 ist eine Dosis-Effekt-Kurve und zeigt die Proliferationsaktivitätsdaten
von Schwann-Zellen, die von der Verabreichung von Aliquots mit unterschiedlicher
Größe von konditioniertem
Medium von CHO-Zellen abstammen.
-
48 ist eine Dosis-Effekt-Kurve bei Schwann-Zellen
und zeigt die mitogenetische Aktivität, die in das extrazelluläre Medium
von SF9 Insektenzellen abgegeben wurde, welche mit Bacculovirus
infiziert waren, die den GGF2HBS5 cDNA-Klon enthielten.
-
49 ist ein Western Blot des konditioniertem Mediums
von rekombinanten CHO-Zellen unter Verwendung eines GGF Peptidantikörpers.
-
50(A) ist eine graphische Darstellung
für Schwann-Zellen
und zeigt die Proliferationsaktivität des rekombinanten (von COS-Zellen
produzierten), menschlichen GGF-II (rhGGF-II) Peaks, der aus der
Kationenaustauschersäule
eluierte. (B) ist ein Immunoblot gegen den rekombinanten GGFII Peak
unter Verwendung des polyklonalen Antikörpers, der gegen das spezifische
Peptid von rhGGFII gemacht wurde.
-
51(A) ist ein Diagramm und zeigt die Reinigung
von rhGGF-II (von CHO-Zellen produziert) auf einer Kationenaustauschersäule mittels
Fraktionierung. (B–C)
sind Fotographien eines Western Blots, der unter Verwendung der
Fraktionen von (A) und eines rhGGF-II spezifischen Antikörpers erstellt
wurde.
-
52 ist die Fotographie eines Gels und veranschaulicht
die Tyrosinphosphorilierung in Schwann-Zellen, die mit rekombinanten
Gliawachstumsfaktoren behandelt wurden.
-
53 zeigt die Sequenzen der GGFHBS5-, GGFHFB1-
und GGFBPP5-Polypeptide
(SEQ ID Nrn. 170, 171 bzw. 172).
-
54 ist eine Karte des CHO-Zell-Expresionsvektors
pcDHFRpolyA.
-
Die
Erfindung bezieht sich auf die Isolierung und Reinigung von neuen
Gliawachstumsfaktoren und auf die Klonierung von DNA-Sequenzen,
welche für
diese Faktoren codieren. Andere Komponenten der Erfindung sind verschiedene
Genspleißvarianten,
die potentiell eine Reihe von Gliawachtumsfaktoren codieren, insbesondere
GGF2HBS5, insbesondere eine Variante, die für das menschliche Äquivalent
von Rind GGF-II codiert. Es ist offensichtlich, dass das Gen, welches
für die
GGF's und p185erbB2 Bindungsproteine codiert, eine Reihe
von unterschiedlich großen,
verschieden gespleißten
RNA-Transkripte produziert, die zu einer Reihe von Proteinen führen, die
eine unterschiedliche Länge
besitzen und einige gemeinsame Peptidsequenzen und einige einzigartige
Peptidsequenzen enthalten. Dies wird unterstützt durch unterschiedlich gespleißte Sequenzen,
die gewonnen werden aus der RNA des Hypophysenhinterlappens vom
Rind (wie hier dargestellt), menschlichem Brustkrebs (MDA-MB-231)
(Holmes et al. Science 256: 1205 (1992) und RNA vom Hühnchengehirn
(Falls et al. Cell 72: 1–20
(1993)). Eine weitere Unterstützung
kommt von Proteinen mit einem breiten Größenbereich, die sowohl als
Mitogene für
Schwann-Zellen (wie hier offenbart) als auch als Liganden für den p185erbB2 Rezeptor wirken (siehe unten).
-
Weitere
Unterstützung
für die
Tatsache, dass die Gene, welche für GGF und p185erbB2 codieren,
homolog sind, kommt von einem Vergleich der Nukleotidsequenzen.
Science, 256 (1992), 1205–1210,
Holmes et al. zeigen die Reinigung eines 45 kD humanen Proteins
(Heregulin-α),
das spezifisch interagiert mit dem Rezeptorprotein p185erbB2,
welches mit verschiedenen menschlichen Bösartigkeiten verbunden ist.
Mehrere komplementäre
DNA-Klone, die für Heregulin-α codieren,
wurden isoliert. Peles et al. (Cell 69: 205 (1992)) und Wen et al
(Cell 69: 559 (1992)) beschreiben eine komplementäre DNA,
die aus Rattenzellen isoliert wurde und für ein Protein codiert, das „neu differentiation
factor" (NDF) genannt
wurde. Das Translationsprodukt von der NDF cDNA hat eine p185erbB2 Bindungsaktivität. Usdin und Fischbach, J.
Cell. Biol. 103: 493–507
(1986); Falls et al., Cold Spring Harbor Symp. Quant. Biol. 55:
397–406
(1990); Harris et al., Proc. Natl. Acad. Sci. USA 88: 7664–7668 (1991)
und Falls et al., Cell 72: 801–815
(1993) zeigen die Reinigung eines 42 kD Glycoproteins, das mit einem
Rezeptorprotein p185erbB2 interagiert und
verschiedene komplementäre
cDNA-Klone wurden isoliert (Falls et al., Cell 72: 801–815 (1993).
Verschiedene andere Gruppen haben die Reinigung von Proteinen mit
verschiedenen Molekulargewichten mit p185erbB2 Bindungsaktivität berichtet.
Diese Gruppen umfassen Lupu et al. (1992), Proc. Natl. Acad. Sci.
USA 89: 2287; Yarden und Peles (1991), Biochemistry 30: 3543; Lupu et
al. (1990), Science 249: 1552; Dobashi et al. (1991), Biochem. Biophys.
Res. Comm. 179: 1536, und Huang et al. (1992), J. Biol. Chem. 257:
11508–11512.
-
Andere Ausführungsformen:
-
Die
Erfindung umfasst jedes Protein, welches im Wesentlichen homolog
zu den Codierungssegmenten der 31 (wie
im folgenden definiert) sowie zu anderen, natürlich vorkommenden GGF-Polypeptiden
ist. Auch umfasst sind allelische Variationen, natürliche Mutanten,
induzierte Mutanten, Proteine, die von DNA codiert werden, die unter
hoch oder niedrig stringenten Bedingungen mit einer Nukleinsäure hybridisieren,
die natürlich
vorkommt (für
Definitionen einer hohen und niedrigen Stringens, siehe Current
Protocols in Molecular Biology, John Wiley & Sons, New York, 1989, 6.3.1–6.3.6),
und Polypeptide oder Proteine, die spezifisch durch Antiseren an
GGF-Polypeptid binden. Der Ausdruck umfasst auch chimäre Polypeptide,
welche die GGF-Polypeptide
umfassen, welche die Sequenzen der 31 umfassen.
-
Die
folgenden Beispiele sollen nicht die Erfindung begrenzen, sondern
sollen die Erfindung erläutern und
eine spezifische Anleitung für
wirksame Präparationstechniken
bereitstellen.
-
Wie
bei dem folgenden Beispiel 3 zu erkennen sein wird, zeigen die vorliegenden
Faktoren eine mitogenetische Aktivität für eine Reihe von Zelltypen.
Die Aktivität
in Bezug auf Fibroplasten zeigt eine Wundreparaturfähigkeit
an und die Erfindung umfasst diese Verwendung. Die allgemeinen Aussagen
zu der Erfindung in Bezug auf Formulierungen und/oder Medikamente
und ihre Herstellung sollen deutlich geeignete Produkte und Verwendungen
umfassen. Dies ist klar eine angemessene Erwartung für die vorliegende
Erfindung unter Betracht von Berichten von ähnlichen Aktivitäten für Fibroplastwachstumsfaktoren
(FGFs). Es kann zum Beispiel verwiesen werden auf Sporn et al., „Peptide
Growth Factors and their Receptors I", Seite 396 (Baird und Bohlen) in dem
Abschnitt mit der Überschrift „FGFs in
Wound Heating and Tissue Repair".
-
Beispiel 1
-
Reinigung von GGF-I und
GGF-II aus Rinderhypophysenhinterlappen
-
I. Herstellung der Faktor-CM
Fraktion
-
4.000
gefrorene ganze Rinderhypophysenhinterlappen (ca. 12 Kilo) wurden über Nacht
aufgetaut, kurz mit Wasser gewaschen und dann mit einem gleichen
Volumen von 0,15 M Ammoniumsulfat in Chargen in einem Waring-Mischer homogenisiert.
Das Homogenat wurde mit 1,0 M HCl auf einen pH von 4,5 eingestellt und
bei 4.900 g für
80 Minuten zentrifugiert. Jegliches fettiges Material in dem Überstand
wurde durch eine Passage durch Glaswolle entfernt. Nach Einstellung
des pH's des Überstandes
auf 6,5 unter Verwendung von 1,0 M NaOH wurde festes Ammoniumsulfat
zugegeben, um eine 36% gesättigte
Lösung
zu erzielen. Es wurde mehrere Stunden gerührt und die Suspension wurde
dann bei 4.900 g für
80 Minuten zentrifugiert und der Niederschlag wurde verworfen. Nach
Filtration durch Glaswolle wurde weiteres festes Ammoniumsulfat
dem Überstand
zugegeben, um eine 75% gesättigte
Lösung
zu erzielen, die dann einmal bei 4.900 g für 80 Minuten zentrifugiert
wurde, nachdem zuvor mehrere Stunden gerührt wurde. Das Pellet wurde
in ca. 2 L von 0,1 M Natriumphosphat, pH 6, 0, resuspendiert und
gegen 3 × 40
L des gleichen Puffers dialysiert. Nach Bestätigung, dass die Leitfähigkeit
des Dialysats unterhalb von 20,0 μSiemens
lag, wurde das Dialysat auf eine Bioprocess-Säule geladen (120 × 113 mm,
Pharmacia), die mit einer Fließrate
von 2 ml pro Minute mit Carboxymethylcellulose (CM-52, Whatman)
bepackt worden war. Die Säule
wurde mit 2 Volumen 0,1 M Natriumphosphat, pH 6,0, gewaschen und
anschließend
mit 2 Volumen von 50 mM NaCl und abschließend mit 2 Volumen von 0,2
M NaCl, beides im gleichen Puffer. Während des abschließendes Schrittes
wurden 10 ml (5 Minuten) Fraktionen gesammelt. Die Fraktionen 73
bis 118 wurden zusammengegeben, gegen 10 Volumen von 10 mM Natriumphosphat,
pH 6,0, zweimal dialysiert und durch Zentrifugation bei 100.000
g für 60
Minuten geklärt.
-
II. Hydroxylapatit-HPLC
-
Hydroxylapaptit-HPLC
war bisher keine Technik, die bei der Isolierung von Gliawachstumsfaktoren verwendet
wurde. Bei der vorliegenden Erfindung hat sie sich jedoch als besonders
effizient herausgestellt. Das von der obigen CM-Cellutosechromatographie erzielte Material
wurde durch einen 0,22 μm
Filter (Nalgene) filtriert, bei Raumtemperatur auf eine Hochleistungshydroxylapatit-Säule (50 × 50 mm, Biorad) geladen, die mit
einer Vorsäule
(15 × 25
mm, Biorad) ausgerüstet
und mit 10 mM Kaliumphosphat, pH 6,0, äquilibriert wurde. Die Elution
bei Raumtemperatur wurde mit einer Fließrate von 2 ml pro Minute unter
Verwendung des folgenden, programmierten, linearen Gradienten durchgeführt:
| Zeit
(Min) | %B |
| 0,0 | 0 |
| 5,0 | 0 |
| 7,0 | 20 |
| 70,0 | 20 |
| 150,0 | 100 |
| 180,0 | 100 |
| 185,0 | 0 |
- Lösungsmittel
A: 10 mM Kaliumphosphat, pH 6,0
- Lösungsmittel
B: 1,0 M Kaliumphosphat, pH 6,0
-
6,0
ml (3 Minuten) Fraktionen wurden während der Gradientenelution
gesammelt. Die Fraktionen 39–45
wurden zusammengefasst und gegen 10 Volumen von 50 mM Natriumphosphat,
pH 6,0, dialysiert.
-
III. Mono S FPLC
-
Mono
S FPLC ermöglicht
ein stärker
konzentrierteres Material für
die nachfolgende Gelfiltration herzustellen.
-
Jedes
partikuläre
Material in dem zusammengefassten Material aus der Hydroxylapatitsäule wurde durch
Klärzentrifugation
bei 100.000 g für
60 Minuten entfernt vor der Beladung auf eine präparative HR10/10 Mono S Kationenaustauschersäule (100 × 10 mm,
Pharmacia), die dann wieder äquilibriert
wurde auf 50 mM Natriumphosphat, pH 6,0, bei Raumtemperatur mit
einer Fließrate
von 1,0 ml/Minute. Unter diesen Bedingungen wurde gebundenes Protein
unter Verwendung des folgenden progammierten linearen Gradienten
eluiert:
| Zeit
(Min) | %B |
| 0,0 | 0 |
| 70,0 | 30 |
| 240,0 | 100 |
| 250,0 | 100 |
| 260,0 | 0 |
- Lösungsmittel
A: 50 mM Kaliumphosphat, pH 6,0
- Lösungsmittel
B: 1,2 M Natriumchlorid, 50 mm Natriumphosphat, pH 6,0
-
1
ml (1 Minute) Fraktionen wurden während diesem Gradientenprogramm
gesammelt. Die Fraktionen 99 bis 115 wurden zusammengefasst.
-
IV. Gelfiltration FPLC
-
Mit
diesem Schritt beginnt die Trennung der zwei Faktoren gemäß der Erfindung
vor der abschließenden
Reinigung und liefert angereicherte Fraktionen.
-
Für die Zwecke
von diesem Schritt wurde eine präparative
Superose 12 FPLC-Säule
(510 × 20
mm, Pharmacia) gemäß den Anleitungen
des Herstellers gepackt. Um diese Säule zu standardisieren, wurde
eine Messung der theoretischen Trennstufen gemäß den Anleitungen des Herstellers
durchgeführt
und es ergab sich ein Wert von 9.700 theoretischen Trennstufen.
-
Das
zusammengefasste Material von dem Mono S eluiertem Material wurde
bei Raumtemperatur in 2,5 ml Aliquots auf dieser Säule in 50
mM Natriumphosphat, 0,75 NaCl, pH 6,0 aufgebracht (zuvor durch eine C18
Umkehrphasensäule
(Sep-pak, Millipore) passiert) bei einer Fließrate von 1,0 ml/Minute. 1
ml (0,5 Minuten) Fraktionen wurden nach 35 Minuten, nachdem jede
Probe auf die Säule
aufgetragen worden war, gesammelt. Die Fraktionen 27 bis 41 (GGF-II)
und 42 bis 57 (GGF-I) wurden bei jedem Lauf zusammengefasst.
-
V. HPLC mit umgekehrten
Phasen
-
Die
GGF-I und GGF-II Pools von den obigen Superose 12 Läufen wurden
jeweils in drei gleiche Aliquots aufgeteilt. Jedes Aliquot wurde
auf eine C8 Umkehrphasensäule
(Aquapore RP-300 7 μ C8
220 × 4,6 mm,
Applied Biosystems) gebracht, geschützt durch eine Vorpatrone (RP-8,
15 × 3,2
mm, Applied Biosystems) und äquilibriert
auf 40°C
mit 0,5 ml pro Minute. Das Protein wurde unter diesen Bedingungen
und unter Verwendung des folgenden, programmierten, linearen Gradienten
eluiert:
| Zeit
(Min) | %B |
| 0 | |
| 60 | 66,6 |
| 62,0 | 100 |
| 72,0 | 100 |
| 75,0 | 0 |
- Lösungsmittel
A: 0,1% Trifluoressigsäure
(TFA)
- Lösungsmittel
B: 90% Acetonitril, 0,1% TFA
-
200 μl (0,4 Minuten)
Fraktionen wurden in silikonisierte Röhrchen (aus „Multilube
tubes", Bioquote)
von 15,2 Minuten nach Beginn des programmierten Gradienten gesammelt.
-
VI. SDS-Polyacrylamid-Gelelektrophorese
-
Bei
diesem Schritt wurden Proteinmolekulargewichtsstandards, niedriger
Bereich, Katalog Nr. 161-0304, von Bio-Rad Laboratories Limited,
Watford, England, verwendet. Die tatsächlich verwendeten Proteine
und ihre Molekulargewichtsstandards wurden zuvor schon angegeben.
-
Die
Fraktionen 47 bis 53 (GGF-I) und die Fraktionen 61 bis 67 (GGF-II)
von den Umkehrphasenläufen wurden
individuell zusammengefasst. 7 μl
des zusammengefassten Materials wurden in einem gleichen Volumen
von 0,0125 M Tris Cl, 4% SDS, 20% Glycerin und 10% β-Mercaptoethanol
für GGF-I
für 5 Minuten
gekocht und auf ein 11% Polyacrylamid-Laemmli-Gel mit einem 4% Stacking-Gel
geladen, und der Lauf erfolgte bei einer konstanten Spannung von
50 V für
16 Stunden. Dieses Gel wurde dann fixiert und gefärbt unter
Verwendung eines Silberfärbekits
(Amersham). Unter diesen Bedingungen wurden die Faktoren jeweils
als eine etwas diffuse Bande bei relativen Molekulargewichten von
30.000 bis 36.000 Daltons (GGF-I) sowie bei 55.000 bis 63.000 Daltons
(GGF-II) gemäß der Definition
durch die Molekulargewichtsmarker erkannt. Aus der Gelfärbung war
abzuleiten, dass es noch eine kleine Zahl an anderen Proteinspezies
mit äquivalenten
Mengen zu den GGF-I und GGF-II Spezies in dem aus den Umkehrphasenläufen zusammengefassten
Material gab.
-
VII. Stabilität in Trifluoressigsäure
-
Die
Stabilitätsdaten
wurden für
die vorliegenden Faktoren in Gegenwart von Trifluoressigsäure wie folgt
erzielt:
GGF-I: Das Material von der Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen wurde in Gegenwart von 0,1% TFA und Acetonitril
innerhalb von 12 Stunden nach Fertigstellung des Säulenlaufs
und dann nach 10-wöchiger
Inkubation bei 40°C überprüft. Nach
der Inkubation hatte GGF-I wenigstens 50% der Aktivität des Materials,
welches direkt nach der Säule
getestet wurde.
GGF-II: Das Material von der Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen wurde in Gegenwart von 0,1% TFA und Acetonitril,
und gelagert bei –20°C, nach dem
Auftauen und dann nach 4-tägiger Inkubation
bei 40°C
getestet. Nach der Inkubation zeigte GGF-II wenigstens 50% der Aktivität des Materials, welches
frisch aufgetaut worden war.
-
Es
ist klar, dass die bei den obigen Studien verwendete Trifluoressigsäurekonzentration
die am meisten verwendete für
die Umkehrphasenchromatographie ist.
-
VIII. Bedingungen für den Aktivitätstest
-
Soweit
nichts anderes angegeben ist, wurden alle Schritte bei 37°C und unter
Verweis auf die 1 bis 6 wurde
die Aktivität
bei jeder Stufe gemäß den Brockes
(Meth. Enz., siehe oben)-Techniken mit den folgenden Modifikationen
bestimmt. Bei der Präparation
der Schwann-Zellen wurde 5 μM
Forskolin zusätzlich zu
der Zugabe von DMEM (Sulbecco's
modifiziertes Eagle's
Medium), FCS und GGF zugesetzt. Die bei dem Test verwendeten Zellen
waren fibroplastenfreie Schwann-Zellen mit einer Passagezahl von
weniger als 10. Diese Zellen wurden aus den Flaschen mit Trypsin
entfernt und in Platten mit einem flachen Boden und mit 96 Vertiefungen
angeordnet zu 3,3 tausend Zellen pro Mikrovertiefung.
-
[125I]ludR wurde für die abschließenden 24
Stunden nach der Testlösungszugabe
zugesetzt. Die (nicht stimulierte) Hintergrundeinlagerung bei jedem
Test betrug weniger als 100 cpm und die maximale Einlagerung war
20- bis 200-fach über
diesem Hintergrund in Abhängigkeit
von der Schwann-Zellcharge
und der Passagezahl.
-
Im
Falle der GGF-I und GGF-II Fraktionen von der oben beschriebenen
Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen wurden zwei Dosis-Effekt-Kurven für jeden
Faktor erstellt, wobei exakt das gleiche Verfahren für jede Kurve
und für
jeden Faktor verwendet wurde. Das obige Verfahren wurde bei dem
Testaufbau nur durch Substitution des fötalen Kalbplasmas für fötales Kalbsserum
modifiziert, um die andere Kurve für jeden Faktor zu erzielen.
Die Ergebnisse sind in den 7 und 8 dargestellt.
-
Beispiel 2
-
Aminosäuresequenzen der gereinigten
GGF-I und GGF-II
-
Die
Studien für
die Analyse der Aminosäuresequenz
wurden durchgeführt
unter Verwendung von hoch gereinigtem Rinderhypophysen GGF-I und
GGF-II. Der konventionelle Einbuchstabencode wurde verwendet, um
die Sequenzen darzustellen. Die Peptide wurden durch Lysylendopeptidase-
und Protease V8-Verdau
erzielt, durchgeführt
mit reduzierten und carboxymethylierten Proben, wobei der Lysylendopeptidase-Verdau
von GGF-II mit Material durchgeführt
wurde, das von der 55–65
RD Region eines 11% SDS-PAGE eluiert wurde (Molekulargewicht relativ
zu den oben angegebenen Markern).
-
Insgesamt
wurden 21 Peptidsequenzen (siehe 9, SEQ
ID Nrn. 1–20,
169) für
GGF-I erzielt, von denen 12 Peptide (siehe 10,
SEQ ID Nrn. 1, 22–29,
17, 19 und 32) nicht in den gegenwärtigen Proteindatenbanken vorhanden
sind und somit einmalige Sequenzen darstellen. Insgesamt 12 Peptidsequenzen
(siehe 11, SEQ ID Nrn. 33–39, 51,
52 und 164–166)
wurden für
GGF-II erzielt, wobei davon 10 Peptide (12, SEQ
ID Nrn. 45–53)
in den gegenwärtigen
Proteindatenbanken nicht vorhanden sind und somit einmalige Sequenzen
darstellen (eine Ausnahme ist das Peptid GGF-II 06, das identische
Sequenzen in vielen Proteinen zeigt, die wahrscheinlich keine Bedeutung
unter Anbetracht der kleinen Zahl an Resten besitzen). Die neuen Sequenzen
entsprechen höchstwahrscheinlich
den Abschnitten von wirklichen Aminosäuresequenzen von GGFs I und
II.
-
Besondere
Aufmerksamkeit sei auf die Sequenzen GGF-I 07 und GGF-II 12 gerichtet,
die sehr stark miteinander verwandt sind. Diese Ähnlichkeiten deuten an, dass
die Sequenzen von diesen Peptiden fast sicher diejenigen der zugeordneten
GGF-Spezies sind, wobei es sehr unwahrscheinlich ist, dass sie von
kontaminierenden Proteinen abstammen.
-
In
dem Peptid GGF-II 02 ist die Sequenz X S S konsistent mit der Gegenwart
einer N-verbundenen Kohlenhydratgruppe auf einem Asparagin bei der
als X bezeichneten Position.
-
In
den
9 und
11 stellt
X im Allgemeinen einen unbekannten Rest dar und kennzeichnet einen Sequenzzyklus,
wo eine einzelne Position nicht mit Sicherheit benannt werden konnte,
weil entweder mehr als ein Signal von gleicher Größe in dem
Zyklus vorlag oder weil kein Signal vorhanden war. Mit Sternchen
sind solche Peptide gekennzeichnet, wo die zuletzt genannte Aminosäure der
letzten Aminosäure
entspricht, die in diesem Peptid vorhanden ist. In den restlichen
Peptiden war die Signalstärke
nach der zuletzt genannten Aminosäure nicht ausreichend, um die
Sequenzbenennung bis zum Ende von diesem Peptid weiterzuführen. Die Säule zur
rechten Hand zeigt die Ergebnisse einer Computerdatenbankrecherche
unter Verwendung der GCG FASTA und TFASTA Paketprogramme, um die
NBRF- und EMBL-Sequenzdatenbanken
zu analysieren. Der Name eines Proteins in dieser Säule bezeichnet
die Identität
eines Teils von seiner Sequenz mit der Peptidaminosäuresequenz,
wobei maximal zwei Fehlanpassungen erlaubt waren. Die verwendeten
Abkürzungen
sind die folgenden:
| HMG-1 | Hochmobilitätsgruppe
Protein-1 |
| HMG-2 | Hochmobilitätsgruppe
Protein-2 |
| LH-alpha | Alpha-Untereinheit
des luteinisierenden Hormons |
| LH-beta | Beta-Untereinheit
des luteinisierenden Hormons |
-
Beispiel 3
-
Mitogenetische Aktivität von gereinigtem
GGF-I und GGF-II
-
Die
mitogenetische Aktivität
von hochgereinigten Proben, die GGFs I und II enthielten, wurden
unter Verwendung einer quantitativen Methode studiert, die es ermöglicht,
dass eine einzelne Mikrokultur hinsichtlich der DNA-Synthese, der Zellmorphologie,
der Zellzahl und der Expression der Zellantigene getestet wird.
Diese Technik wurde modifiziert von einem Verfahren, das zuvor berichtet
wurde von Muir et al., Analytical Biochemistry 185, 377–382, 1990.
Die Hauptmodifikationen waren: 1) die Verwendung von nicht-beschichteten
Mikrotiterplatten, 2) die Zellzahl pro Vertiefung, 3) die Verwendung
von 5% fötalem
Rinderplasma (FBP) anstelle von 10% fötalem Kalbsserum (FCS), und
4) die Zeit der Inkubation in Gegenwart des Mitogens und Bromdeoxyuridin
(BrdU), die simultan den Kulturen zugesetzt wurden. Ferner wurde
die Zellmonoschicht vor der Fixierung nicht gewaschen, um einen
Verlust an Zellen zu vermeiden. Die Inkubationszeit des monoklonalen Maus
Anti-BrdU Antikörpers
und des Peroxidase-konjugierten Ziege Anti-Maus Immunoglobulin (IgG)
Antikörpers
wurden verdoppelt, um die Empfindlichkeit des Tests zu erhöhen. Der
Test, der für
Schwann-Zellen des Schiasnervs der Ratte optimiert war, wurde auch
für verschiedene
Zelllinien nach geeigneter Modifikation der Zellkulturbedingungen
verwendet.
-
I. Verfahren der Mitogenesetestung
-
Am
Tag 1 wurden gereinigte Schwann-Zellen auf nicht-beschichtete Platten
mit 96 Vertiefungen in 5% FBP/Dulbecco's Modified Eagle Medium (DMEM) (5.000
Zellen/Vertiefung) aufgetragen. Am Tag 2 wurden GGFs und andere
Testfaktoren den Kulturen zugegeben, sowie BrdU mit einer Endkonzentration
von 10 μm. Nach
48 Stunden (Tag 4) wurde die BrdU-Einlagerung durch Absaugen des Mediums
beendet und die Zellen wurden mit 200 μl/Vertiefung von 70% Ethanol
für 20
Minuten bei Raumtemperatur fixiert. Die Zellen wurden dann mit Wasser
gewaschen und die DNA wurde durch Inkubation mit 100 μl 2 N HCl
für 10
Minuten bei 37°C denaturiert.
Nach Absaugung wurde die Restsäure
durch Füllen
der Vertiefungen mit 0,1 M Boratpuffer, pH 9,0, neutralisiert und
die Zellen wurden mit Phosphat gepufferter Kochsalzlösung (PBS)
gewaschen. Die Zellen wurden dann mit 50 μl Blockpuffer (PBS, 0,1% Triton × 100 und
2% normales Ziegenserum enthaltend) für 15 Minuten bei 37°C behandelt.
Nach Absaugung wurde der monoklonale Maus Anti-BrdU Antikörper (Dako Corp.,
Santa Barabara, CA) (50 μl/Vertiefung;
1,4 μg/ml,
verdünnt
in Blockpuffer) zugegeben und für
2 Stunden bei 37°C
inkubiert. Nicht gebundene Antikörper
wurden durch drei Waschungen in PBS entfernt. Mit Peroxidase konjugierter
Ziege Anti-Maus IgG Antikörper
(Dako Corp., Santa Barbara, CA) (50 μl/Vertiefung; 2 μg/ml, verdünnt in Blockpuffer)
wurde zugegeben und für
1 Stunde bei 37°C
inkubiert. Nach dreimaligem Waschen in PBS/Triton und einer abschließenden Spülung in
PBS wurden 100 μl/Vertiefung
von 50 mM Phosphat/Citrat-Puffer, pH 5,0, in die Vertiefungen gegeben,
wobei der Puffer 0,05% an löslichem
Chromogen o-Phenylendiamin (OPD) und 0,02% H2O2 enthielt. Die Reaktion wurde nach 5–20 Minuten
bei Raumtemperatur durch Pipettieren von 80 μl von jeder Vertiefung zu einer
sauberen Platte beendet, die 40 μl/Vertiefung
an 2 N Schwefelsäure
enthielt. Die Extinktion wurde bei 490 nm unter Verwendung eines
Plattenlesers (Dynatech Labs) bestimmt. Die Testplatten, welche
die Zellmonoschichten enthielten, wurden zweimal mit PBS gewaschen
und immunozytochemisch gefärbt
für BrdU-DNA,
indem 100 μl/Vertiefung
des Substrates Diaminobenzidin (DAB) und 0,02% H2O2 hinzugegeben wurde, um ein nicht lösliches
Produkt zu bilden. Nach 10–20
Minuten wurde die Farbreaktion durch Waschen mit Wasser gewaschen
und die BrdU-positiven Kerne wurden unter Verwendung eines Umkehrmikroskops
beobachtet und gezählt.
Gelegentlich wurden negative Kerne mit 0,001% Toluidinblau gegengefärbt und
wie oben gezählt.
-
II. Zelllinien, die für die Mitogenesetests
verwendet wurden
-
Swiss
3T3 Fibroplasten: Zellen von Flow Labs wurden in DMEM, ergänzt mit
10% FCS, Penicillin und Streptomycin bei 37°C in einer feuchten Atmosphäre von 10%
CO2 in Luft gehalten. Die Zellen wurden
gefüttert oder
alle zwei Tage subkultiviert. Für
den mitogenetischen Test wurden die Zellen mit einer Dichte von
5.000 Zellen/Vertiefung in Komplettmedium gegeben und für 1 Woche
inkubiert, bis die Zellen zusammenfließend und ruhig waren. Das Medium
enthaltende Serum wurde entfernt und die Zellmonoschicht wurde zweimal
mit Serum freiem Medium gewaschen. 100 μl von Serum freiem Medium, das
die Mitogene und 10 μM
an BrdU enthielt, wurden zu jeder Vertiefung gegeben und für 48 Stunden
inkubiert. Die Dosisreaktionen auf GGFs und Serum oder PDGF (als
eine positive Kontrolle) wurden aufgenommen.
-
BHK
(Babyhamsternieren) 21 C13 Fibroplasten: Die Zellen von European
Coltection of Animal Cell Cultures (ECACC), wurden gehalten in Glasgow
Modified Eagle Medium (GMEM), das mit 5% Tryptosephosphatnährtösung, 5%
FCS, Penicillin und Streptomycin ergänzt war. Die Zellen wurden
bei 37°C
in feuchter Atmosphäre
von 5% CO2 in Luft gehalten. Die Zellen
wurden gefüttert
oder alle 2 bis 3 Tage subkultiviert. Für den mitogenetischen Test
wurden die Zellen mit einer Dichte von 2.000 Zellen/Vertiefung in
einem Komplettmedium für
24 Stunden angeordnet. Das Serum enthaltende Medium wurde dann entfernt
und es wurde mit Serum freiem Medium gewaschen und ersetzt mit 100 μl von 0,1%
FCS, das GMEM enthielt, oder GMEM alleine. GGFs und FCS oder bFGF
als positive Kontrollen wurden zugegeben, zusammen mit 10 μM BrdU, und für 48 Stunden
inkubiert. Die Zellkulturen wurden dann verarbeitet, wie oben für die Schwann-Zellen
beschrieben.
-
C6
Ratten Glioma-Zelllinie: Zellen, die bei der Passage 39 erzielt
wurden, wurden in DMEM, das 5% FCS, 5% Pferdeserum (HS), Penicillin
und Streptomycin enthielt, bei 37°C
in einer feuchten Atmosphäre
von 10% CO2 in Luft gehalten. Die Zellen
wurden gefüttert
oder alle 3 Tage subkultiviert. Für den mitogenetischen Test
wurden die Platten mit einer Dichte von 2.000 Zellen/Vertiefung
in einem Komplettmedium angeordnet und für 24 Stunden inkubiert. Dann
wurde das Medium nach Waschung in Serum freiem Medium ersetzt durch einer
Mischung von 1 : 1 DMEM und F12 Medium, das 0,1% FCS enthielt. Die
Dosisreaktionen auf GGFs, FCS und αFGF wurden dann durchgeführt und
die Zellen wurden mittels ELISA verarbeitet, wie dies zuvor für die anderen
Zelltypen beschrieben worden war.
-
PC12
(Rattennebennieren-Pheochromocytomazellen): Zellen von ECACC wurden
in RPMI 1640, ergänzt
mit 10% HS, 5% FCS, Penicillin und Streptomycin, in Kollagen beschichteten
Flaschen bei 37°C
in einer feuchten Atmosphäre
von 5 CO2 in Luft gehalten. Die Zellen wurden
gefüttert
alte 3 Tage durch Ersatz von 80% des Mediums. Für den mitogenetischen Test
wurden die Zellen mit einer Dichte von 3.000 Zellen/Vertiefung in Komplettmedium
auf Kollagen beschichteten Platten angeordnet (50 μl/Vertiefung
Kollagen, Vitrogen Collagen Corp., verdünnt 1 : 50, 30 Minuten bei
37°C) und
für 24
Stunden inkubiert. Das Medium wurde dann ersetzt durch frisches
RPMI, entweder alleine oder 1 mM Insulin oder 1% FCS enthaltend.
Dosisreaktionen auf FCS/HS (1 : 2) als positive Kontrolle und auf
GGFs wurden wie oben durchgeführt.
Nach 48 Stunden wurden die Zellen fixiert und ELISA wurde, wie oben
beschrieben, durchgeführt.
-
III. Ergebnisse der mitogenetischen
Tests
-
Alle
Experimente in diesem Beispiel wurden unter Verwendung einer hoch
gereinigten Probe aus dem Sepharose 12 Chromatographiereinigungsschritt
(siehe Beispiel 1, Abschnitt D), die eine Mischung von GGF-I und
GGF-II (GGFs) enthielt, durchgeführt.
-
Zunächst wurden
die Ergebnisse, die mit dem BrdU Einlagerungstest erzielt wurden,
mit dem klassischen mitogenetischen Test für Schwann-Zellen verglichen,
der basiert auf einer [125] I-UdR Einlagerung in DNA von sich teilenden
Zellen, wie dies beschrieben ist von J. P. Brockes (Methods Enzymol.
147: 217, 1987).
-
13 zeigt den Vergleich der mit den beiden Tests
erzielten Daten, wobei die gleichen Zellkulturbedingungen (5.000
Zellen/Vertiefung in 5% FBP/DMEM; inkubiert in Gegenwart von GGFs
für 48
Stunden) angelegt wurden. Wie deutlich zu erkennen ist, sind die
Ergebnisse vergleichbar, wobei der BrdU-Einlagerungstest etwas mehr
empfindlich erscheint, wie dies durch die Verlagerung der Kurve
zur linken Seite der graphischen Darstellung erscheint, das heißt, zu niedrigeren
Konzentrationen von GGFs.
-
Wie
unter dem Abschnitt „Verfahren
für die
Mitogenesetestung" beschrieben,
können
die Originaltestplatten, welche die Zellmonoschichten enthalten,
die zweite Reaktion durchführen,
was zu dem unlöslichen DAB-Produkt führt, welches
die BrdU positiven Kerne erbt, nachdem die immunoreaktive BrdU-DNA
durch Lesen der Intensität
des löslichen
Produktes der OPD Peroxidasereaktion quantifiziert worden ist. Die
Mikrokulturen können
dann unter einem Umkehrmikroskop überprüft werden und die Zellmorphologie
und die Zahl der BrdU positiven und der negativen Kerne können beobachtet
werden.
-
In
der 14a und in der 14b ist die BrdU-DNA Immunoreaktivität, die durch
Ablesen der Extinktion bei 490 nm bestimmt wurde, verglichen mit
der Zahl der BrdU positiven Kerne und mit dem Prozentsatz der BrdU
positiven Kerne hinsichtlich der Gesamtzahl der Zellen pro Vertiefung,
was in den gleichen Kulturen gezählt
wurde. Die Standardabweichungen waren weniger als 10%. Die zwei
Prüfverfahren
zeigen eine gute Korrelation und die Diskrepanz zwischen den Werten
bei der höchsten
Dosis der GGFs kann durch das unterschiedliche Maß der DNA-Synthese
in den Zellen, die als BrdU positiv detektiert wurden, erklärt werden.
-
Der
BrdU Einlagerungstest kann somit nützliche zusätzliche Information über die
biologische Aktivität der
Polypeptide auf die Schwann-Zellen bereitstellen, wenn der Test
mit dem (125) I-UdR Einlagerungstest verglichen wird. Die in der 15 dargestellten Daten zeigen, dass GGFs auf die
Schwann-Zellen wirken kann, um die DNA-Synthese zu induzieren, wobei
aber bei niedrigeren Dosen die Zahl der negativen Zellen, die in der
Mikrokultur nach 48 Stunden vorhanden sind, zunimmt.
-
Der
Test wurde dann auf verschiedene Zelllinien von unterschiedlichem
Ursprung angewandt. In der 16 sind
die mitogenetischen Reaktionen der Schwann-Zellen und der Swiss
3T3 Fibroplasten auf GGFs verglichen. Trotz der schwachen Antwort
bei den 3T3 Fibroplasten wurden in diesen Kulturen einige deutliche BrdU
positive Kerne nachgewiesen. Kontrollkulturen liefen parallel in
Gegenwart von unterschiedlichen Dosierungen von FCS oder menschlichem
rekombinanten PDGF, was anzeigt, dass die Zellen in der Lage waren, auf
geeignete Anreize zu reagieren (nicht gezeigt).
-
Die
Fähigkeit
der Fibroplasten zur Reaktion auf GGFs wurde ferner untersucht unter
Verwendung der BHK 21 C13 Zelllinie. Diese Fibroplasten, die von
der Niere abstammten, zeigten keine Kontakthemmung und erreichten
ein Ruhestadium, wenn sie zusammen geflossen waren. Deshalb wurden
die experimentellen Bedingungen so gewählt, dass eine sehr geringe
Hintergrundproliferation vorhanden war, ohne dass die Zellvitalität beeinträchtigt wurde.
GGFs besitzen eine signifikante mitogenetische Aktivität auf die
BHK 21 C13 Zellen, wie dies in den 17 und 18 dargestellt
ist. Die 17 zeigt die BrdU Einlagerung
in die DNA durch BHK 21 C13 Zellen, welche durch GGFs in Gegenwart
von 0,1% FCS stimuliert war.
-
Die
gute mitogenetische Reaktion auf FCS zeigt, dass die Zellkulturbedingungen
nicht limitierend waren. In der 18 ist
der mitogenetische Effekt von GGFs als Zahl der BrdU positiven Zellen
und der BrdU negativen Zellen und als die Gesamtzahl der Zellen,
die pro Vertiefung gezählt
wurden, ausgedrückt.
Die Daten sind repräsentativ
für zwei
experimentelle Läufe
in zweifacher Ausfertigung, wobei wenigstens drei Felder pro Vertiefung
gezählt
wurden. Wie bei Schwann-Zellen beobachtet, führt GGFs zusätzlich zu
einem proliferativen Effekt bei niedrigen Dosen auch zu einer Erhöhung der
Zahl der überlebenden
Zellen, die nicht reagieren. Der Prozentsatz der BrdU positiven
Zellen ist proportional zu der Zugabemenge von GGFs, das den Kulturen
zugesetzt wurde. Die Gesamtzahl der Zellen nach 48 Stunden in Gegenwart
von höheren
Dosen von GGFs ist wenigstens verdoppelt, was bestätigt, dass
GGFs die DNA-Synthese und die Proliferation in den BHK 21 C13 Zellen
induziert. Unter den gleichen Bedingungen zeigten Zellen, die für 48 Stunden
in Gegenwart von 2% FCS gehalten wurden, eine Zunahme um das 6-fache
(nicht gezeigt).
-
C6
Gliomazellen sind ein nützliches
Modell zum Studium der Eigenschaften von Gliazellen. Der exprimierte
Phenotyp scheint abhängig
von der Zellpassage zu sein. Zu einem frühen Stadium ähneln die
Zellen mehr dem Astrozytenphänotyp,
während
ein Oligodentrozytenphänotyp
zu späteren
Stadien auftritt (jenseits von Passage 70). C6 Zellen, die bei diesen
Experimenten verwendet wurden, stammten von der Passage 39 bis Passage
52. C6 Zellen sind eine hoch proliferierende Population, so dass
die experimentellen Bedingungen so optimiert wurden, dass nur ein
sehr geringer Hintergrund an BrdU Einbau auftrat. Die Gegenwart
von 0,1% Serum war notwendig, um die Zellvitalität zu erhalten, ohne dass dies
signifikant die mitogenetischen Reaktionen beeinflusste, wie dies
durch die Dosisreaktion auf FCS (19)
dargestellt ist.
-
In
der 20 sind die mitogenetischen Reaktionen
auf aFGF (saurer Fibroplastenwachstumsfaktor) und auf GGFs als Prozentsätze des
maximalen BrdU Einbaus dargestellt, der in Gegenwart von FCS (8%)
erzielt wurde. Die Werte sind Durchschnittswerte von zwei Experimenten
in zweifacher Ausfertigung. Die Wirkung der GGFs war vergleichbar
mit der einer reinen Zubereitung von aFGF. AFGF wurde als ein spezifischer Wachstumsfaktor
für C6
Zellen beschrieben (Lim R. et al., Cell Regulation 1: 741–746, 1990)
und deshalb wurden sie als positive Kontrolle hier verwendet. Das
direkte Zählen
der BrdU positiven und negativen Zellen war nicht möglich, da
in den Mikrokulturen eine hohe Zelldichte vorlag. Im Gegensatz zu
den bisher berichteten Zelllinien zeigten PC12 Zellen keine deutliche
Reaktion auf GGFs, wenn sie unter Kulturbedingungen behandelt wurden,
bei denen PC12 auf Seren reagieren kann (Mischung von FCS und HS,
wie dies routinemäßig für die Zellerhaltung
verwendet wird). Die Zahl der pro Vertiefung vorhandenen Zellen
scheint jedoch das Verhalten der PC12 Zellen zu beeinflussen, so
dass weitere Experimente erforderlich sind.
-
Beispiel 4
-
Isolierung und Klonierung
von Nukleotidsequenzen, die für
Proteine codieren, welche GGF-I und GGF-II Peptide enthalten
-
Die
Isolierung und die Klonierung der GGF-II Nukleotidsequenzen wurde
durchgegeführt,
wie hier dargestellt, unter Verwendung der Peptidsequenzinformation
und Selektion aus einer Bibliothek. Diese wurde durchgeführt, wie
unten dargestellt. Es ist deutlich, dass die Peptide der 4 und 5 als
Ausgangspunkt für
die Isolierung und Klonierung der GGF-I Sequenzen gemäß den hier
beschriebenen Techniken verwendet werden können. In der Tat zeigt die 21, SEQ ID Nrn. 54–76 und 78–88 mögliche degenerierte Oligonukleotidproben
für diesen
Zweck und die 23(A–B), SEQ ID Nrn. 90–119, listet
mögliche
PCR Primer auf. Die DNA-Sequenz und die Polypeptid-Sequenz sollte
mit diesen Mitteln erhältlich
sein, wie bei GGF-II,
und auch die DNA-Konstrukte und die Expressionsvektoren, welche
solche DNA-Sequenzen aufnehmen, Wirtszellen, die genetisch verändert sind
durch Einbau von solchen Konstrukten/Vektoren sowie Proteine, die
durch die Kultivierung von solchen Wirtszellen erzielbar sind. Die
Erfindung umfasst solche Gegenstände.
-
I. Aufbau und Synthese
von Oligonukleotidsonden und Primer
-
Degenerierte
DNA-Oligomersonden wurden durch Rücktranslation der Aminosäuresequenzen
(abstammend von den Peptiden, die von dem gereinigten GGF-Protein
erzeugt wurden) in Nukleotidsequenzen erstellt. Die Oligomere stellten
entweder den codierenden Strang oder den nicht-codierenden Strang der DNA-Sequenz dar.
Wenn Serin, Arginin oder Leucin in dem Oligomeraufbau vorhanden
waren, wurden zwei getrennte Synthesen durchgeführt, um Mehrdeutigkeiten zu
vermeiden. Serin wird zum Beispiel durch TCN oder durch AGY, wie
bei 537 und 539 oder bei 609 und 610, codiert. Eine ähnliche
Codonaufteilung wurde für Arginin
oder Leucin vorgenommen (zum Beispiel 544, 545). Die DNA-Oligomere
wurden auf einem Biosearch 8750 4-Säulen DNA-Synthesegerät unter
Verwendung der β-Cyanoethylchemie
und betrieben auf der 0,2 Mikromol-Syntheseskala, synthetisiert.
Die Oligomere wurden von der Säule
(500 Angstrom CpG Harze) abgespalten und in konzentriertem Ammoniumhydroxid
für 6–24 Stunden
bei 55–60°C entschützt. Die
entschützten Oligomere
wurden unter Vakuum (Speedvac) getrocknet und mittels Elektrophorese
in Gelen von 15% Acrylamid (20 Mono 1 Bis) gereinigt mit 50 mM Tris-Borat-EDTA
Puffer, der 7 M Harnstoff enthielt. Die Oligomere mit voller Länge wurden
in Gelen durch UV-Abschattung nachgewiesen und dann wurden die Banden
ausgeschnitten und die DNA-Oligomere
wurden unter Schütteln
in 1,5 ml H2O für 4 bis 16 Stunden eluiert.
Das Eluat wurde getrocknet, in 0,1 ml H2O
wieder gelöst
und bei 260 nm wurde die Extinktion gemessen.
-
Die
Konzentrationen wurden gemäß der folgenden
Formel bestimmt: (A 260 × Einheiten/ml)(60,6/Länge = × μM)
-
Durch
Zugabe von H2O wurden alle Oligomere auf
50 μM Konzentration
eingestellt.
-
Die
wie oben aufgebauten, degenerierten Sonden sind in der 21, SEQ ID Nrn. 54–76 und 78–88, gezeigt.
-
Die
PCR-Primer wurden im Wesentlichen mit den gleichen Verfahren hergestellt,
die auch für
die Sonden verwendet wurden, wobei aber die folgenden Modifikationen
eingeführt
wurden. Linker von dreizehn Nukleotiden, die Restriktionsstellen
enthielten, wurden an den 5' Enden
der degenerierten Oligomere, um die Restriktionsstellen beim Klonieren
in Vektoren zu benutzen. Die DNA-Synthese wurde auf der 1 Mikromol-Skala unter
Verwendung von 1.000 Angstrom CpG Harzen durchgeführt und
Inosin wurde an den Positionen verwendet, wo alte vier Nukleotide
normal in die degenerierten Proben eingeführt wurden. Die Reinigung der
PCR-Primer umfasste eine Ethanolfällung mit anschließender Gelelektrophoresereinigung.
-
II. Aufbau der Bibliothek
und Screening
-
Eine
genomische Rinder-DNA-Bibliothek wurde von Stratagene (Katalog Nr.
945701) gekauft. Die Bibliothek enthielt 2 × 106 15–20 kb Sau3A1
partielle Rinder-DNA-Fragmente, die in den Vektor Lambda DashII kloniert
waren. Eine cDNA-Bibliothek von dem gesamten Rindergehirn wurde
gekauft von Clonetech (Katalog Nr. BL 10139). Komplementäre DNA-Bibliotheken
wurden von mRNA hergestellt (In Vitrogen; Stratagene), wobei die
mRNA aus dem Rindergesamthirn, der Rinderhypophyse und dem Rinderhypophysenhinterlappen
hergestellt wurde. In Vitrogen stellte zwei cDNA-Bibliotheken her:
Eine Bibliothek war in dem Vektor Lambda g10 und die andere Bibliothek
war in dem Vektor pcDNAI (eine Plasmidbibliothek). Die Stratagene-Bibliotheken wurden
in dem Vektor Lambda unizap hergestellt.
-
Zusammenfassend
enthielten die cDNA-Bibliotheken 14 Millionen primär rekombinante
Phagen.
-
Die
genomische Rinderbibliothek wurde auf E. Coli K12 Wirtsstamm LE392
auf 23 × 23
cm Platten (Nunc) mit 150.000 bis 200.000 Phagenplaques pro Platte
aufgetragen. Jede Platte stellte etwa ein genomisches Rinderäquivalent
dar. Nach einer über
Nacht Inkubation bei 37°C
wurden die Platten abgekühlt
und Replikafilter wurden gemäß dem Verfahren
von Maniatis et al. (2: 60–81)
hergestellt. Vier Plaquelifts wurden von jeder Platte auf nicht-beladenen
Nylonmembranen hergestellt (Pall Biodyne A oder MSI Nitropure).
Die DNA wurde auf den Membranen durch Quervernetzung unter UV-Licht
für 5 Minuten
oder durch Backen bei 80°C unter
Vakuum für
2 Stunden immobilisiert. Die DNA-Sonden wurden unter Verwendung
von T4-Polynukleotidkinase (New England Biotabs) mit gamma-32P ATP
(New England Nuclear; 6500 Ci/mMol) gemäß den Angaben des Lieferanten
markiert. 50 pMole von degenerierten DNA-Oligomeren wurden in Gegenwart
von 600 μCi gamma 32P-ATP und 5 Einheiten der T4-Polynukleotidkinase
für 30
Minuten bei 37°C
inkubiert. Die Reaktionen wurden beendet, der Ladepuffer für die Gelelektrophorese
wurde hinzugefügt
und die radiomarkierten Sonden wurden mit Elektrophorese gereinigt.
Die 32P markierten Sonden wurden aus den Gelstückchen ausgeschnitten und im
Wasser eluiert. Alternativ wurden die DNA-Sonden über die
PCR-Amplifizierung durch Einlagerung von α-32P-dATP oder α-32P-dCTP
markiert gemäß dem Protokoll
von Schowalter und Sommer, Anal. Biochem 177: 90–94 (1989). Die in den PCR-Reaktionen
markierten Sonden wurden mittels Entsalzung auf Sephadex G-150 Säulen gereinigt.
-
Die
Prehybridisierung und die Hybridisierung wurden durchgeführt in GMC
Puffer (0,52 M NaPi, 7% SDS, 1% BSA, 1,5 mM EDTA, 0,1 M NaCl, 10
mg/ml tRNA). Das Waschen wurde durchgeführt in Oligowaschungen (160
ml 1 M Na2HPO4,
200 ml 20% SDS, 8,0 ml 0,5 M EDTA, 100 ml 5 M NaCl, 3632 ml H2O). Typischerweise wurden 20 Filter (jeweils
400 cm2), die Replikatkopien von zehn Rindergenomäquivalenten
darstellten, in 200 ml Hybridisierungslösung mit 100 pMole degenerierter
Oligonukleotidsonde (128–512
fach degeneriert) inkubiert. Die Hybridisierung geschah über Nacht
mit 5°C
unterhalb der Minimumschmelztemperatur, die für die degenerierte Sonde berechnet
wurde. Die Berechnung der Minimumschmelztemperatur basiert auf 2°C für ein AT-Paar und auf 4°C für ein GC-Paar.
-
Die
Filter wurden unter wiederholtem Wechseln der Oligowaschungen bei
den Hybridisierungstemperaturen für 4 bis 5 Stunden gewaschen
und abschließend
in 3,2 M Tetramethylammoniumchlorid, 1% SDS, zweimal für 30 Minuten
bei einer Temperatur in Abhängigkeit
von der DNA-Sondenlänge.
Für 20mers
betrug die abschließende
Waschtemperatur 60°C.
Die Filter wurden aufgelegt und dann wurde mit Röntgenstrahlfilm (Kodak XAR5)
unter Verwendung von Verstärkerfolien
(Dupont Cronex Lightening Plus) entwickelt. Normalerweise war eine
drei- bis fünftägige Filmentwicklung
bei –80°C ausreichend,
um die Duplikatsignale in diesen Bibliothekstests zu detektieren.
Nach Analyse der Ergebnisse konnten die Filter abgezogen und wieder
verwendet werden. Die Filter wurden abgezogen durch Inkubation über zwei
aufeinander folgende Zyklen von 15 Minuten in einem Mikrowellenofen
mit voller Kraft in einer Lösung
von 1% SDS, das 10 mM EDTA pH 8, enthielt. Die Filter wurden wenigstens
für drei
bis vier Zyklen von Stripping und wieder beladen mit verschiedenen Sonden
verwendet.
-
III. Isolierung und Wachstum
von rekombinanten Phagen und DNA-Aufarbeitung
-
Diese
Verfahren wurden gemäß dem Standardprotokoll
durchgeführt,
das beschrieben ist in „Recombinant
DNA" (Maniatis et
al. 2: 60–2:
81).
-
IV. Analyse der isolierten
Klone unter Verwendung von DNA-Verdau und Southern Blots
-
Rekombinante
Phagen-DNA-Proben (2 Mikrogramm) wurden gemäß den Bedingungen geschnitten, die
von dem Lieferanten der Restriktionsendonuklease empfohlen waren
(New England Biolabs). Nach einer vierstündigen Inkubation bei 37°C wurden
die Reaktionsprodukte in Gegenwart von 0,1 M Natriumacetat und drei
Volumen Ethanol gefällt.
Die gefällte
DNA wurde mittels Zentrifugation gesammelt, in 75% Ethanol gespült und dann
getrocknet. Alle resuspendierten Proben wurden auf Agarosegels aufgetragen
(typischerweise 1% in TAE-Puffer; 0,04 M Tris-Acetat; 0,002 M EDTA).
Die Gele liefen bei 1 Volt pro Zentimeter für 4 bis 20 Stunden. Die Marker
umfassten lambda Hind III DNA-Fragmente und/oder ∅X174HaeIII
DNA-Fragmente (New
England Biolabs). Die Gele wurden mit 0,5 Mikrogramm/ml von Ethidiumbromid
gefärbt
und fotografiert. Für
das Southern Blotting wurde die DNA in dem Gel durch Behandlung
mit 0,125 N HCl depuriniert, in 0,5 N NaOH denaturiert und in 20 × SSC überführt (3 M
Natriumchlorid; 0,03 M Natriumcitrat), um die Nylonmembranen zu entladen.
Das Blotting wurde über
einen Zeitraum von 6 bis 24 Stunden durchgeführt und dann wurden die Filter
in 0,5 Tris-HCL, pH 7,5, 0,15 M Natriumchlorid neutralisiert und
dann kurz in 50 mM Tris-Borat-EDTA gespült.
-
Für die Vernetzung
wurden die Filter zuerst in eine transparente Kunststofffolie eingewickelt
und die DNA-Seite wurde dann für
5 Minuten ultraviolettem Licht ausgesetzt. Die Hybridisierung und
das Waschen wurde durchgeführt,
wie dies für
die Bibliothekstestung (siehe Abschnitt 2 von diesem Beispiel) beschrieben ist.
Für die
Hybridisierungsanalyse zur Bestimmung, wo ähnliche Gene in anderen Arten
existieren, wurden leichte Modifikationen durchgeführt. Der
DNA-Filter wurde gekauft von Clonetech (Katalog Nr. 7753-1) und
enthielt 5 Mikrogramm von EcoRI geschnittene DNA von verschiedenen
Arten pro Linie. Die Sonde wurde mit den PCR-Amplifikationsreaktionen markiert, wie
dies oben im Abschnitt 2 beschrieben ist. Die Hybridisierungen wurden
durchgeführt
in 80% Puffer B (2 g Polyvinylpyrrolidin, 2 g Ficoll-400, 2 g Rinderserumalbumin,
50 ml 1 M Tris-HCl (pH 7,5) 58 g NaCl, 1 g Natriumpyrophosphat,
10 g Natriumdodecylsulfat, 950 ml H2O),
der 10% Dextransulfat enthielt. Die Sonden wurden durch Kochen für 10 Minuten
und anschließendes
schnelles Abkühlen
in Eiswasser denaturiert. Die Sonde wurde dem Hybridisierungspuffer
mit 106 dpm 32P pro ml zugegeben und über Nacht bei 60°C inkubiert.
Die Filter wurden bei 60°C
zunächst
gewaschen in Puffer B, gefolgt von 2 × SSC, 0,1% SDS, dann in 1 × SSC, 0,1%
SDS. Für
Experimente mit hoher Stringens wurden die abschließenden Waschungen
in 0,1 × SSC,
1% SDS durchgeführt
und die Temperatur wurde auf 65°C
angehoben.
-
Die
Ergebnisse der Southern Blots wurden verwendet, um eine Restriktionskarte
des genomischen Klons herzustellen und um darzustellen, welche Subfragmente
mit den GGF-Sonden (Kandidaten für
die Subklonierung) hybridisierten.
-
V. Subklonierung der DNA-Segmente,
die zu den Hybridisierungssonden homolog waren
-
Die
DNA-Verdaus (zum Beispiel 5 Mikrogramm) wurden auf 1% Agarosegels
aufgebracht und geeignete Fragmente wurden nach Anfärbung aus
den Gelen geschnitten. Die DNA wurde durch Adsorption an Glaskügelchen
und anschließender
Elution unter Verwendung des Protokolls, wie es von dem Lieferanten
(Bio 101) beschrieben war, gereinigt. Die gewonnenen DNA-Fragmente (100–200 ng)
wurden in linearisierte, dephosphorilierte Vektoren, zum Beispiel
pT3T7 (Ambion), was ein Derivat von pUC18 ist, unter Verwendung
der T4-Ligase (New England Biolabs) ligiert. Dieser Vektor trägt das E.
Coli β-Lactamasegen,
so dass die Transformanten auf Platten mit Ampicillin selektiert
werden können.
Der Vektor führt
auch zu einer β-Galactosidase-Komplementierung
der Wirtszelle, so dass nicht-Rekombinanten (blau) unter Verwendung
von Isopropylthiogalactosid und Bluogal (Bethesda Research Labs)
delektiert werden konnten. Ein Teil der Ligierungsreaktionen wurde
verwendet für
die Transformation von E. Coli K12 XL1 blau-kompetente Zellen (Stratagene,
Katalog Nr. 200236). Die Transformanten wurden dann auf LB-Platten
selektiert, die 50 Mikrogramm pro ml Ampicillin enthielten. Weiße Kolonien
wurden ausgewählt
und Plasmid-mini-Präparationen
wurden für
den DNA-Verdau und für
die DNA-Sequenzanalyse hergestellt. Die ausgewählten Klone wurden wieder getestet,
um zu bestimmen, ob ihre insertierte DNA mit den GGF-Sonden hybridisierte.
-
VI. DNA-Seguenzierung
-
Doppelsträngige Plasmid-DNA-Template
wurden hergestellt aus 5 ml Kulturen gemäß Standardprotokollen. Die
Sequenzierung wurde durch die Dideoxy-Kettenabbruchreaktion unter
Verwendung von Sequenase 2.0 und einem Dideoxynukleotid-Sequenzierkits
(US Biochemical) gemäß dem Protokoll
der Hersteller durchgeführt
(eine Modifikation von Sanger et al. PNAS; USA 74: 5463 (1977)).
Alternativ wurde die Sequenzierung in einem thermischen DNA-Zyklussequenzierer
(Perkin Elmer, Modell 4800) unter Verwendung eines Zyklussequenzierkits
(New England Biolabs; Bethesda Research Laboratories) durchgeführt, wobei
die Angaben des Herstellers unter Verwendung eines 5'-endmarkierten Primers angewandt wurden.
Die Sequenzprimer waren solche, die mit den Sequenzierkits geliefert
wurden, oder wurden gemäß der Sequenz
synthetisiert, die von den Klonen bestimmt wurde. Die Sequenzierreaktionsansätze wurden
auf ein 0,4 mm dickes Sequenziergel von 6% Polyacrylamid geladen
und dort aufgelöst.
Die Gele wurden getrocknet und mit Röntgenstrahlfilm entwickelt.
Typischerweise wurde 35S eingelagert, wenn Standardsequenzierkits
verwendet wurden, und 32P endmarkierte Primer wurden verwendet für die Zyklussequenzierreaktionen.
Die Sequenzen wurden in einem DNA-Sequenziereditor vom Boden der
Gele bis zu ihrer Spitze (5' zu
3' Richtung) gelesen
und die Daten wurden analysiert unter Verwendung von Programmen,
die von Genetics Computer Group (GCG, University of Wisconsin) geliefert
wurden.
-
VII. RNA-Zubereitung und
PCR-Amplifizierung
-
Offene
Leserahmen, die in der genomischen DNA entdeckt wurden und die Sequenzen
für die
Kodierung von GGF-Peptiden enthielten, wurden via PCR-Amplifizierung von
Hypophysen-RNA verlängert.
Die RNA wurde hergestellt aus gefrorenem Rindergewebe (Pelfreeze)
gemäß dem Guanidin
neutal-CsCl Verfahren (Chirgwin et. al. Biochemistry 18: 5294 (1979).
Polyadenylierte RNA wurde ausgewählt
mit Oligo-dT Cellulosesäulenchromatographie
(Aviv und Leder, PNAS (USA) 69: 1408 (1972)).
-
Spezifische
DNA-Zielsequenzen wurden amplifiziert, wobei entweder mit Gesamt-RNA
begonnen wurde oder mit polyadenylierten RNA-Proben, die zu cDNA
unter Verwendung des Perkin Elmer PCR/RNA Kits Nr.: N808-0017 umgewandelt
worden waren. Die strangreversen Transkriptionsreaktionen verwendeten
1 μg Templat-RNA
und Primer von Oligo-dT mit angehefteten Linkern mit Restriktionsenzymstellen,
oder spezifische Antisense-Primer mit angehefteten Restriktionsorten,
wobei die Primer von klonierten Sequenzen bestimmt wurden. Um den
zweiten Strang herzustellen, waren die Primer entweder einmalige
Plusstrangsequenzen, wie sie verwendet wurden bei den 3' RACE Reaktionen
(Frohman et. al., PNAS (USA) 85: 8998 (1988)), oder Oligo-dT Primer mit angebrachten
Restriktionsstellen, wenn die zweite Zielstelle durch terminales
Transferasetailing an die ersten Strangreaktionsprodukte mit dATP
hinzugefügt
worden ist (z. B. 5' RACE
Reaktion, Frohman et. al., ibid). Alternativ, wie bei anchored-PCR-Reaktionen,
wurden die zweiten Strangprimer degeneriert und repräsentierten
somit besondere Peptidsequenzen.
-
Das
Amplifizierungsprofil folgte dem folgenden allgemeinen Schema: 1)
Einweichen des Ansatzes für 5
Minuten bei 95°C;
2) thermaler Zyklus Ansätze
von 1 Minute, 95°C;
1 Minute abflachen auf eine Schmelztemperatur von 45°C, 50°C oder 55°C; Aufrechterhaltung
der Schmelztemperatur für
1 Minute; Erwärmen über 1 Minute
auf 72°C;
Ausdehnen bei 72°C
für 1 Minute
oder für
1 Minute plus eine 10 Sekunden Autoausdehnung; 3) Ausdehnungszyklus
bei 72°C,
5 Minuten und 4) Einweichen des Ansatzes bei 4°C für unbegrenzte Zeit. Die thermischen
Zyklusansätze
(Nr. 2) liefen normalerweise für
30 Zyklen. Eine sechzehn μl
Probe von jeder 100 μl
Amplifizierungsreaktion wurde durch Elektrophorese in 2% Nusieve
1% Agarose Gels analysiert, welche in TAE-Puffer bei 4 Volt pro Zentimeter für 3 Stunden
tiefen. Die Gele wurden gefärbt
und dann gegen nicht beladene Nylonmembranen geblottet, welche mit
markierten DNA-Sonden, die intern zu den Primern waren, getestet.
-
Spezifische
Sets der DNA-Amplifizierungsprodukte konnten bei den Blottingexperimenten
identifiziert werden und ihre Positionen wurden als Anhaltspunkt
für die
Reinigung und Reamplifikation verwendet. Wo es geeignet war, wurden
die restlichen Teile der ausgewählten
Proben auf präparative
Gele geladen und nach Elektrophorese wurden vier bis fünf Stücke von
0,5 mm Dicke (welche die erwartete Position des spezifischen Produktes
umfassten) dem Gel entnommen. Die Agarose wurde zerdrückt und
dann in 0,5 ml Elektrophoresepuffer bei 40°C für 2–16 Stunden eingeweicht. Die zerdrückte Agarose
wurde für
2 Minuten zentrifugiert und die wässrige Phase wurde in frische
Röhrchen
transferiert.
-
Die
Reamplifikation wurde auf fünf
Mikroliter (etwa 1% des Produktes) des eluierten Materials durchgeführt, wobei
die gleichen Sets von Primer und Reaktionsprofilen, wie bei den
Originalreaktionen, verwendet wurden. Nach Vollendung der Reamplifikationsreaktionen
wurden die Proben mit Chloroform extrahiert und in frische Röhrchen transferiert.
Konzentrierte Restriktionsenzympuffer und Enzyme wurden den Reaktionen
zugesetzt, um sie an den Restriktionsstellen, die in den Linkern
vorhanden waren, zu schneiden. Die verdauten PCR-Produkte wurden
mit Gelelektrophorese gereinigt und dann in Vektoren subkloniert,
wie dies oben in dem Subklonierungsabschnitt beschrieben ist. Die
DNA-Sequenzierung wurde wie oben beschrieben, durchgeführt.
-
VIII. DNA-Sequenzanalyse
-
Die
DNA-Sequenzen wurden unter Verwendung eines Fragmentassemblierprogramms
bereitgestellt und die Aminosäuresequenzen
wurden unter Verwendung der GCG-Programme GelAssemble, Map und Translate
abgeleitet. Die abgeleiteten Proteinsequenzen wurden verwendet als
Abfragesequenz, um Proteinsequenzdatenbanken unter Verwendung von
WordSearch zu durchsuchen. Die Analyse wurde auf einer VAX Station
3100 Workstation, die unter VMS 5.1 lief, gemacht. Die Datenbankrecherche
wurde gemacht auf SwissProt Freigabenummer 21 unter Verwendung der
GCG Version 7,0.
-
IX. Ergebnisse der Klonierung
und der Sequenzierung von Genen, die für GGF-I und GGF-II codieren
-
Wie
oben dargelegt wurden zur Identifikation der DNA-Sequenz, die für Rind GGF-II
codiert, degenerierte Oligonukleotidsonden aus GGF-II Peptidsequenzen
erstellt. GGF-II 12 (SEQ ID Nr. 44), ein Peptid, welches via Lysylendopeptidaseverdau
einer gereinigten GGF-II Zubereitung (siehe die 11 und 12)
erstellt wurde, zeigte eine starke Aminosäuresequenzhomologie mit GGF-I
07 (SEQ ID Nr. 39), ein tryptisches Peptid, das aus einer gereinigten
GGF-I Zubereitung hergestellt wurde. GGF-II 12 wurde somit verwendet,
um zehn degenerierte Oligonukleotidproben zu erzeugen (siehe Oligos
609, 610 und 649 bis 656 in der 21, SEQ
ID Nrn. 69, 70, 71 und 79). Ein zweifacher Satz an Filtern wurden
mit zwei Sätzen
(Satz 1 = 609, 610; Satz 2 = 649–5656) von Sonden getestet,
die für
zwei überlappende
Abschnitte von GGF-II 12 codieren. Hybridisierungssignale wurden
beobachtet, wobei aber nur ein Klon mit beiden Sondensätzen hybridisierte.
Der Klon (als GGF2BG1 bezeichnet) wurde gereinigt.
-
Die
Southern Blot Analyse der DNA aus dem Phargenklon GGF2BG1 bestätigte, dass
beide Sätze
der Sonden mit dieser Rind-DNA-Sequenz hybridisierten und zeigte
ferner, dass beide Sonden mit dem gleichen Satz an DNA-Fragmenten
innerhalb des Klons reagierte. Basierend auf diesen Experimenten
wurde ein 4 kb Eco RI Subfragment des Ursprungsklons identifiziert,
subkloniert und partiell sequenziert. Die 22 zeigt
die Nukleotidsequenz (SEQ ID Nr. 89) und die abgeleitete Aminosäuresequenz
(SEQ ID Nr. 196) der anfänglichen DNA-Sequenzlesungen,
welche die Hybridisierungsstellen der Sonden 609 und 650 umfassten,
und bestätigte,
dass ein Teil dieser genomischen Rind DNA für das Peptid 12 (KASLADSGEYM;
SEQ ID Nr. 129) codiert.
-
Die
weitere Sequenzanalyse zeigte, dass GGF-II 12 auf einem offenen
Leserahmen von 66 Aminosäuren
beruht (siehe unten), was der Startpunkt wurde für die Isolierung der überlappenden
Sequenzen, die ein mutmaßliches
GGF-II Rindergen und eine cDNA darstellen.
-
Verschiedene
PCR-Verfahren wurden verwendet, um zusätzliche Codiersequenzen für das mutmaßliche GGF-II
Rindergen zu erhalten. Gesamt-RNA
und Oligo dT-ausgewählte
(Poly A enthaltend) RNA Proben wurden aus der Rindergesamthypophyse,
des Hypophysenvorderlappens, des Hypophysenhinterlappens und aus
dem Hypothalamus hergestellt. Unter Verwendung der Primer aus der
Liste, die in der 23 gezeigt ist, SEQ
ID Nrn. 109–119,
wurden einseitige PCR-Reaktionen (RACE) verwendet, um die cDNA-Enden
in der 3' sowie
in der 5' Richtung
zu amplifizieren. Anchored PCR-Reaktionen wurden mit den degenerierten
Oligonukleotidprimern durchgeführt,
welche zusätzliche
GGF-II Peptide darstellen. Die 24 fasst
die angrenzenden DNA-Strukturen und die Sequenzen, die bei diesen
Experimenten erzielt wurden, zusammen. Von den 3' RACE-Reaktionen wurden drei alternativ
gespleißte
cDNA-Sequenzen hergestellt, die kloniert und sequenziert wurden.
Eine 5' RACE-Reaktion
führte
zur Enddeckung eines zusätzlichen
Exons, das die codierende Sequenz für wenigstens 52 Aminosäuren enthielt.
Die Analyse der abgeleiteten Aminosäuresequenz ergab die Peptide
GGF-II-6 und eine Sequenz ähnlich
zu GGF-I-18 (siehe unten). Die anchored-PCR-Reaktionen führten zu
der Identifikation der (cDNA) codierenden Sequenzen der Peptide
GGF-II-1, 2, 3 und 10, die innerhalb eines zusätzlichen cDNA-Segmentes von
300 Basenpaaren enthalten waren. Die 5' Grenze dieses Segmentes (das heißt Segment
E, siehe 31) wird durch das Oligonukleotid
definiert, welches für
das Peptid GGF-II-1 codiert und das in der PCR-Reaktion verwendet
wurde (zusätzliche
5' Sequenzdaten
existieren, wie dargestellt, für
den menschlichen Klon von Beispiel 6). Somit enthält dieser
Klon die Nukleotidsequenz, die für
sechs von den insgesamt neun neuen GGF-II Peptidsequenzen codieren.
-
Das
klonierte Gen wurde zunächst
charakterisiert durch die Erstellung einer physikalischen Karte
von GGF2BG1, welche es uns erlaubte, die codierenden Sequenzen,
wie sie gefunden worden waren (siehe unten, 25),
zu positionieren. Die DNA-Sonden von den codierenden Sequenzen,
die oben beschrieben sind, wurden verwendet, um weitere DNA-Fragmente
zu identifizieren, welche die Exons von diesem Phagenklon enthielten,
und um Klone, welche in beiden Richtungen überlappen, zu identifizieren.
Das mutmaßliche
GGF-II Rindergen wird in wenigstens fünf codierende Segmente aufgeteilt.
Die codierenden Segmente werden als diskrete Längen der DNA-Sequenz definiert,
die in Polypeptidsequenzen unter Verwendung des universellen genetischen
Codes translatiert werden können.
Die in der 31 beschriebenen, codierenden
Segmente, auf die in der vorliegenden Anwendung verwiesen wird,
sind: 1) einzelne Exons, die innerhalb des GGF-Gens (zum Beispiel codierendes Segment
a) vorhanden sind oder 2) abstammen von Sätzen von zwei oder mehr Exons,
die in spezifischen Untergruppen der mRNAs erscheinen, wobei jeder
Satz in die spezifischen Polypeptidsegmente, wie in den Genprodukten
gezeigt, translatiert werden können.
Die Polypeptidsegmente, auf die in den Ansprüchen bezogen wird, sind die
Translationsprodukte der analogen codierenden DNA-Segemente. Nur
die codierenden Segmente A und B wurden als Exons definiert und
sequenziert und bisher kartiert. Die Übersicht für die angrenzenden, identifizierten,
codierenden Sequenzen ist in der 26 wiedergegeben.
Die Exons sind aufgelistet (alphabetisch) in der Reihenfolge ihrer
Entdeckung. Es ist von den Intron/Exon-Grenzen erkennbar, das Exon
B in den cDNAs enthalten sein kann, welche das codierende Segment
E und das codierende Segment A verbinden. Dies bedeutet, dass Exon
B nicht herausgespleißt
werden kann, ohne den Leserahmen zu beeinträchtigen. Deshalb vermuten wir,
dass drei alternative Spleißmuster
die mutmaßlichen GGF-II
cDNA-Rindsequenzen 1, 2 und 3 herstellen können. Die codierenden Sequenzen
davon, bezeichnet als GGF2BPP1.CDS, GGF2BPP2.CDS und GGF2BPP3.CDS,
sind dargestellt in den 28A (SEQ
ID Nr. 133), 28B–C (SEQ ID Nr. 134) bzw. 28D–E
(SEQ ID Nr. 135). Die abgeleiteten Aminosäuresequenzen von diesen drei
cDNAs sind auch in den 28A (SEQ
ID Nr. 190), 28B–C (SEQ ID Nr. 191) und 28D–E
(SEQ ID Nr. 143) gezeigt.
-
Die
drei abgeleiteten Strukturen codieren für Proteine mit einer Länge von
206, 281 und 257 Aminosäuren.
Die ersten 183 Reste der abgeleiteten Proteinsequenzen sind in allen
drei Genprodukten identisch. An der Position 184 unterscheiden sich
die Klone signifikant. Ein Codon für Glycin GGT in GGF2BPP1 dient
auch als Spleißdonor
für GGF2BPP2
und GGF2BPP3, der alternativ auf den Exons C, C/D, C/D' und D oder C, C/D bzw.
D hinzugefügt
ist und in der 33 (SEQ ID Nr. 149)
gezeigt ist. GGFIIBPP1 ist ein abgestumpftes Genprodukt, das erzeugt
wird durch Lesen vorbei an der Spleißstelle des codierenden Segmentes
A in die folgende Intron-Sequenz (Intron). Dies stellt das codierende
Segment A' in der 31 dar (SEQ ID Nr. 140). Das Transkript
endet benachbart zu einer kanonischen AATAAA Polyadenylierungssequenz,
wobei wir vermuten, dass dieses abgestumpfte Genprodukt ein echtes
reifes Transkript darstellt. Die zwei anderen, längeren Genprodukte teilen sich
die gleiche 3' nicht
translatierte Sequenz und die Polyadenylierungsstelle.
-
Alle
drei Moleküle
enthalten sechs der neun, neuen GGF-II Peptidsequenzen (siehe 12) und ein anderes Peptid ist hoch homolog zu
GGF-I-18 (siehe 27). Dieses Ergebnis beinhaltet
eine hohe Wahrscheinlichkeit, dass dieses rekombinante Molekül wenigstens
für einen
Teil des Rind GGF-II codiert. Ferner sind die berechneten, isoelektrischen
Punkte für
die drei Peptide konsistent mit den physikalischen Eigenschaften
von GGF-I und II. Da die Molekülgröße von GGF-II
etwa 60 kD beträgt,
sollte die längste
der drei cDNAs für
ein Protein codieren mit annähernd
der Hälfte
der vorausgesagten Zahl an Aminosäuren.
-
Eine
Probe, welche die Exons B und A umfasste, wurde mittels PCR-Amplifizierung markiert
und verwendet, um eine cDNA-Bibliothek zu screenen, welche aus RNA
hergestellt wurde, die aus Rinderhypophysenhinterlappen isoliert
wurde. Ein Klon (GGF2BPP5) zeigte das Muster, das in der 30 angezeigt ist, und enthielt ein zusätzliches
codierendes DNA-Segment (G) zwischen den codierenden Segmenten A
und C. Die gesamte Nukleinsäuresequenz
ist in der 32 gezeigt (SEQ ID Nr.
148). Das vorhergesagte Translationsprodukt von dem längsten offenen
Leserahmen hat 241 Aminosäuren.
Ein Teil von einer zweiten cDNA (GGF2BPP4) wurde auch aus der Bibliothek
des Rinderhypophysenhinterlappens unter Verwendung der oben beschriebenen
Sonde isoliert. Dieser Klon zeigte das Muster, das in der 30 dargestellt ist. Dieser Klon ist an dem 5' Ende nicht vollständig, wobei
er aber eine Spleißvariante
in dem Sinne darstellt, dass die codierenden Segmente G und D fehlen.
BPP4 zeigt auch ein neues 3' Ende
mit den Regionen H, K und L jenseits der Region C/D. Die Sequenz
von BPP4 ist in der 34(A–C) gezeigt
(SEQ ID Nr. 150).
-
Beispiel 5
-
GGF-Sequenzen
in verschiedenen Arten
-
Eine
Datenbankrecherche hat keine bedeutungsvollen Ähnlichkeiten zwischen einem
prognostizierten GGF Translationsprodukt und bekannten Proteinsequenzen
ergeben. Dies lässt
vermuten, dass GGF-II das erste Mitglied einer neuen Familie oder
Superfamilie von Proteinen ist. Bei Kreuzhybridisierungsstudien
mit hoher Stringens (DNA-Blottingversuche) mit anderen Säugetier-DNAs
haben wir klar gezeigt, dass DNA-Sonden von diesem rekombinanten
Rindermolekül
spezifische Sequenzen in einer Reihe von getesteten Proben ohne weiteres
nachweisen kann. Eine hoch homologe Sequenz wurde auch in der menschlichen
genomischen DNA detektiert. Das Autoradiogramm ist in der 29 gezeigt. Die Signale in den Spuren, die Ratten
und menschliche DNA enthielten, stellen die menschlichen und Ratten Äquivalente
des GGF Gens dar, wobei die Sequenzen für die verschiedenen cDNA's, welche für dieses
Gen codieren, vor kurzem dargestellt wurden von Holmes et al., (Science
256: 1205 (1992)) und Wen et al. (Cell 69: 559 (1992)).
-
Beispiel 6
-
Isolierung der menschlichen
Sequenz, die für
das menschliche GGF2 codiert
-
Verschiedene
menschliche Klone, die Sequenzen aus dem GGF-II codierenden Segment
E vom Rind enthielten, wurden durch Screening einer menschlichen
cDNA-Bibliothek isoliert, welche aus dem Gehirnstamm isoliert wurde
(Stratagene Katalog Nr. 935206). Diese Strategie wurde verfolgt
auf Basis der starken Verbindung zwischen den meisten GGF2-Peptiden
(einmalig vorhanden für
GGF2) und der prognostizierten Peptidsequenz aus den Klonen, die
das E-Rindersegment enthielten. Diese Bibliothek wurde, wie im Beispiel 4,
Abschnitt II beschrieben, getestet unter Verwendung der Oligonukleotidsonde
914–919,
die im folgenden aufgeführt
sind.
-
-
Die
mit diesen Sonden nachgewiesenen Klone wurden ferner mittels Hybridisierung
analysiert. Eine von dem codierenden Segment A (siehe 21) abstammende Sonde, welche durch Markierung
eines Polymerasekettenreaktion (PCR)-Produktes vom Segment A hergestellt
wurde, wurde auch verwendet, um die primäre Bibliothek zu testen. Verschiedene
Klone, die mit den von A und E abstammenden Sonden hybridisierten, wurden
ausgewählt
und ein besonderer Klon, GGF2HBS5, wurde für die weitere Analyse ausgewählt. Dieser Klon
ist dargestellt durch das Muster der codierenden Segmente (EBACC/D'D, wie in der 31 gezeigt). Das E-Segment in diesem Klon
ist das menschliche Äquivalent
der abgestumpften Rinderversion von E, die in der 37 gezeigt
ist. GGF2HBS5 ist der wahrscheinlichste Kandidat für die Codierung
von GGF-II von all den „mutmaßlichen" GGF-II Kandidaten.
Die Länge
des codierenden Sequenzsegmentes E beträgt 786 Nukleotide plus 264
Basen einer nicht-translatierten Sequenz. Die prognostizierte Größe des Proteins,
welches von GGF2HBS5 codiert wird, beträgt etwa 423 Aminosäuren (ungefähr 45 Kilodaltons,
siehe 45(A–D), SEQ ID Nr. 170), was in
etwa der Größe der deglykosylierten
Form von GGF-II entspricht (siehe Beispiel 16). Ferner haben sieben
der GGF-II Peptide, die in der 27 aufgeführt sind, äquivalente
Sequenzen, die in die Proteinsequenz, die von der Region E prognostiziert
wurde, fallen. Die Peptide II-6 und II-12 sind Ausnahmen und fallen
in das codierende Segment B bzw. in das codierende Segment A. Die
GGF2HBS5 proteincodierende RNA wurde in einem in vitro Transkriptionssystem
hergestellt, das von dem Bakteriophagen T7-Promotor angetrieben
wird, der in dem Vektor (Bluescript SK (Stratagene Inc.) siehe 44) vorliegt, welcher das GGF2HBS5-Insert enthält. Diese
RNA wurde in einem zellfreien (Kaninchen-Reticulyt) Transtationssystem translatiert
und die Größe des Proteinproduktes
betrug 45 kD.
-
Das
zellfreie Produkt wurde ferner in einem mitogenetischen Schwann-Zelltest überprüft, um die
biologische Aktivität
zu bestätigen.
Die mit konditioniertem Medium behandelten Schwann-Zellen zeigten
eine erhöhte
Proliferation, gemessen durch die Einlagerung von 125I-Uridin,
und eine Phosphorilierung am Tyrosin von einem Protein im 185 Kilodalton-Bereich.
Die Größe des von
GGF2HBS5 codierten Produktes und die Gegenwart der DNA-Sequenzen, welche
für menschliche
Peptide codieren, die hoch homolog zu den Rinderpeptiden sind, gezeigt
in der 12, bestätigen, dass GGF2HBS5 für das menschliche Äquivalent
von Rind-GGF2 codiert. Die Tatsache, dass konditioniertes Medium,
welches aus mit diesem Klon transformierten Zellen hergestellt wurde,
die mitogenetische Aktivität
der Schwann-Zellen auslöst, bestätigt, dass
das GGFIIHBS5-Genprodukt (verschieden von dem BPP5-Genprodukt) sekrediert
wird. Ferner scheint das GGFIIBPP5-Genprodukt die Reaktion der Schwann-Zellproliferation über eine
Rezeptortyrosinkinase, wie p185erbB2, oder
einen nahen verwandten Rezeptor (siehe Beispiel 14) zu vermitteln.
-
Beispiel 7
-
Expression von menschlichem
rekombinanten GGF2 in Säugetierzellen
und Insektenzellen
-
Der
GGF2HBS2 cDNA Klon, der für
das menschliche GGF2 codiert (wie im Beispiel 6 beschrieben und hier
auch als HBS5 bezeichnet) wurde kloniert in den Vektor pcDL-SRα296 (Takebe
et al., Mol. Cell. Biol. 8: 466–472
(1988) und COS-7 Zellen wurden in 100 mm Schalen transfiziert mittels
dem DEAE-Dextranverfahren (Sambrook
et al. Molecular Cloning: A Laboratory Manual, 2. Auflage CSH Laboratory
NY (1989). Zelllysate oder konditioniertes Medium von Transient
exprimierenden COS-Zellen wurden drei oder vier Tage nach der Transfektion
geerntet. Um die Lysate herzustellen, wurden die Zellmonoschichten
mit PBS gewaschen, von den Schalen abgekratzt und durch drei Einfrier/Auftau-Zyklen
in 150 μl
von 0,25 M Tris-HCl, pH 8,0, lysiert. Die Zelltrümmer wurden pelletiert und
der Überstand
wurde verwendet. Proben von konditionierten Medien (7 ml) wurden
gesammelt, dann konzentriert und der Puffer wurde ausgetauscht mit
10 mM Tris, pH 7,4, unter Verwendung von Centiprep-10 und Centricon-10-Einheiten,
wie vom Hersteller beschrieben (Amicon, Beverly, MA). Schwann-Zellen
von Rattennerven wurden hinsichtlich der Einlagerung von DNA-Synthesevorläufern, wie
beschrieben (Beispiel 3), getestet. Proben von konditionierten Medien
oder Zelllysaten wurden in dem Schwann-Zellproliferationstest überprüft, wie
dies in Beispiel 3 beschrieben ist. Die mitogenetischen Aktivitätsdaten
sind in der 46 dargestellt. Die cDNA, GGF2HBS5,
die GGF2 codiert, steuerte die Sekretion es Proteinproduktes in
das Medium. Ein kleiner Teil der Gesamtaktivität war innerhalb der Zellen
nachweisbar, wie dies durch Tests unter Verwendung von Zelllysaten
bestimmt wurde. GGF2HFB1 und GGFBPP5 cDNA's steuerten nicht die Sekretion des
Produktes zu dem extrazellulären
Medium. Die GGF-Aktivität
von diesen Klonen wurde nur in den Zelllysaten nachgewiesen (46).
-
Rekombinantes
GGF2 wurde auch in CHO-Zellen exprimiert. Die GGF2HBS5 cDNA, die
für GGF2
codiert, wurde in die EcoRI-Stelle des Vektors pcdhfrpolyA (54) kloniert und in die DHFR negative CHO-Zelllinie
(DG44) transfiziert mittels des Calciumphosphat-Coprezipitationsverfahrens
(Graham und Van Der Eb, Virology 52: 456–467 (1973). Die Klone wurden
in einem nukleotid- und nukleosidfreien α-Medium (Gibco) in Platten mit
96 Vertiefungen ausgewählt.
Nach 3 Wochen wurden Proben von konditioniertem Medium von unterschiedlichen
Klonen hinsichtlich der Expression von GGF durch den Schwann-Zellproliferationstest,
beschrieben in Beispiel 3, getestet. Stabile Klone, die signifikante
Mengen von GGF-Aktivität
in das Medium abgaben, wurden identifiziert. Die Daten der Schwann-Zellproliferationsaktivität von verschiedenen
Volumenaliquots von CHO-Zellen konditioniertem Medium wurden verwendet,
um die Dosisreaktionskurve aufzustellen, die in der 47 gezeigt ist (Graham und Van Der Eb, Virology
52: 456, 1973). Dieses Material wurde auf einem Western-Blot analysiert,
der mit polyklonalen Antiseren, die gegen ein GGF2 spezifisches
Peptid erstellt wurden, untersucht wurde. Ein breites Band von etwa
69–90
kD (die erwartete Größe von GGF2,
extrahiert von Hypophysen und höher
Molekulargewichts-Glycoformen) wurde spezifisch markiert (49, Spur 12).
-
Rekombinantes
GGF2 wurde auch in Insektenzellen unter Verwendung der Bacculovirusexpression exprimiert.
Sf9 Insektenzellen wurden mit Bacculovirus infiziert, welche den
GGF2HBS5 cDNA Klon enthielten, mit einer Menge von 3–5 (106 Zellen/ml) und in Sf900-II Medium (Gibco)
kultiviert. Die mitogenetische Aktivität der Schwann-Zellen wurde
in das extrazelluläre
Medium (48) abgegeben. Verschiedene
Volumen von konditioniertem Medium der Insektenzellen wurden in
dem Schwann-Zellproliferationstest in Abwesenheit von Forskolin
getestet und die verwendeten Daten zur Herstellung der Dosisreaktionskurve
sind in der 48 gezeigt.
-
Dieses
Material wurde auch auf einem Western-Blot (47)
analysiert, der mit dem oben beschriebenen GGF-II spezifischen Antikörper untersucht
wurde. Ein Band von 45 kD, was der Größe von deglycosiliertem GGF-II
(siehe Beispiel 16) entspricht, wurde gesehen.
-
Die
in diesem Beispiel verwendeten Verfahren waren die folgenden:
Die
mitogenetische Schwann-Zellaktivität von rekombinanten, menschichen
und Rinder-Gliawachstumsfaktoren wurde wie folgt bestimmt:
Die
mitogenetischen Reaktionen von kultivierten Schwann-Zellen wurden
in Gegenwart von 5 μM
Forskolin gemessen, wobei rohe rekombinante GGF-Zubereitungen verwendet wurden, die
von den transienten Säugetierexpressionsexperimenten
erzielt wurden.
-
Einlagerung
von [125I]-Uridin wurde bestimmt nach einer
18 bis 24 Stunden Exposition von Materialien, die von transfizierten
oder scheintransfizierten COS-Zellen, wie beschrieben bei den Verfahren,
erzielt wurden. Die mittlere und die Standardabweichung von vier
Datensätzen
sind dargestellt. Die mitogenetische Reaktion auf die partiell gereinigte,
native Rinderhypophysen GGF (Carboxymethylcellulosefraktion; Goodearl
et al., eingereicht) als Standard von 100% Aktivität dargestellt
(GGF).
-
cDNAs
(53) wurden in pcDL-SRα296 (Takebe et al., Mol. Cell
Biol. 8: 466–472
(1988)) kloniert und COS-7 Zellen wurden in 100 mm Schalen transfiziert
mittels des DEAE-Dextranverfahrens (Sambrook et al., In Motecular
Cloning. A Laboratory Manual, 2. Auflage (Cold Spring Harbor Laboratory
Press, Cold Spring Harbor, NY, 1989)). Zelllysate oder konditionierte
Medien wurden 3 oder 4 Tage nach der Transfektion geerntet. Um die
Lysate herzustellen, wurden die Zellmonoschichten mit PBS gewaschen,
von den Schalen abgekratzt und durch drei Einfrier/Auftau-Zyklen
in 150 μl
von 0,25 M Tris-HCl, pH 8, lysiert. Die Zelltrümmer wurden pelletiert und
der Überstand
wurde verwendet. Proben von konditionierten Medien (7 ml) wurden
gesammelt, dann konzentriert und der Puffer wurde mit 10 mM Tris,
pH 7,4, ausgetauscht unter Verwendung von Centriprep-10 und Centricon-10
Einheiten, wie dies von dem Hersteller beschrieben ist (Amicon,
Beverly, MA). Schwann-Zellen
vom Rattenischiasnerv wurden hinsichtlich der Einlagerung von DNA-Synthesevorläufern getestet,
wie beschrieben von Davis und Stroobant, J. Cell Biol. 110: 1353–1360 (1990);
Brockes et al., Brain Res. 165: 105–118 (1979)).
-
Western-Blots
von konditionierten Medien von rekombinanten CHO-Zellen wurden wie
folgt durchgeführt:
Ein rekombinanter CHO-Klon wurde in 7 ml von MCDB302 proteinfreiem
Medium für
3 Tage kultiviert. 2 ml des konditionierten Mediums wurde konzentriert,
und der Puffer wurde gegen 10 mM Tris-HCl, pH 7,4, ausgetauscht
und bis zur Trockenheit lyophilisiert. Das Pellet wurde in SDS-PAGE
Probenpuffer resuspendiert und einer reduzierenden SDS-Gelelektrophorese
unterworfen und durch Western-Blotting mit einem GGF-Peptidantikörper analysiert.
Eine CHO-Kontrolle wurde unter Verwendung von konditioniertem Medium
von nicht-transfiziertem CHO-DG44 Wirt durchgeführt und die CHOHBS5 Mengen
wurden unter Verwendung von konditioniertem Medium aus einem rekombinanten
Klon getestet.
-
Beispiel 8
-
Isolierung von anderen
menschlichen Sequenzen, die mit Rind-GGF verwandt sind
-
Die
Ergebnisse der Beispiele 5 und 6 zeigen, dass verwandte Sequenzen
von GGF aus menschlichen Quellen auch einfach isoliert werden können unter
Verwendung der DNA-Sonden, die von GGF-Rindersequenzen abstammen.
Alternativ kann das von Holmes et al. (Science 256: 1205 (1992))
beschriebene Verfahren verwendet werden. In diesem Beispiel wurde
ein menschliches Protein (Heregulin α), welches an den p185erbB2 Rezeptor bindet und diesen aktiviert
(und das mit GGF verwandt ist), aus einer Tumorzelllinie gereinigt
und die erhaltende Peptidsequenz wurde verwendet, um Oligonukleotidsonden
herzustellen, die verwendet wurden, um cDNAs, welche für Heregulin
codieren, zu klonieren. Der biochemische Test für die p185erbB2 Rezeptoraktivierung
ist verschieden von der Schwann-Zellproliferation. Dies ist ein ähnliches
Verfahren zu den in den Beispielen 1–4 für die Klonierung der GGF-Sequenzen
aus Hypophysen-cDNAs verwendeten Verfahren. Das Heregulinprotein
und die komplementären
DNAs wurden aus den Tumorzelllinien gemäß den folgenden Verfahren isoliert.
-
Heregulin
wurde aus Medium gereinigt, das durch MDA-MB-231 Brustkrebszellen
konditioniert war (ATCC # HTB 26), gewachsen auf Percell Biolytica
Microcarrier-Beads (Hyclone Labs). Das Medium (10 Liter) wurde 25-fach durch Filtration
durch eine Membran (10 kD Absperrung) (Millipore) konzentriert und
durch Zentrifugation und Filtration durch einen Filter (0,22 μm) gereinigt.
Das Filtrat wurde auf eine Heparin-Sepharose-Säule (Pharmacia) aufgetragen
und die Proteine wurden in Schritten von 0,3, 0,6 und 0,9 M NaCl
in physiologische Kochsalzlösung,
gepuffert mit Phosphat, eluiert. Die Aktivität in den verschiedenen chromatographischen
Fraktionen wurde gemessen durch Quantifizierung der Zunahme bei
der Tyrosinphosphorilierung von p185erbB2 in
MCF-7 Brusttumorzellen (ATCC # HTB 22). MCF-7 Zellen wurden auf
Costar-Platten mit 24 Vertiefungen und F12 (50%) Dulbecco's essentielles Minimalmedium
(50%) gegeben, das Serum (10%) (105 Zellen
pro Vertiefung) enthielt. Zur Festhaftung standen die Zellen für wenigstens
24 Stunden. Vor dem Test wurden die Zellen in ein Medium ohne Serum
für mindestens
1 Stunde überführt. Die
Säulenfraktionen
(10 bis 100 μl)
wurden für
30 Minuten bei 37°C
inkubiert. Die Überstände wurden
dann verdampft und die Reaktion wurde durch Zugabe von SDS-PAGE
Probenpuffer (100 μl)
gestoppt. Die Proben wurden für
5 Minuten auf 100°C erhitzt
und Teile (10 bis 15 μl)
wurden auf ein Tris-Glycingel aufgetragen (4 bis 20%) (Novex). Nach
der Elektrophorese wurden die Proteine auf eine Polyvinylidendifluorid
(PVDF)-Membran elektrogeblottet und dann mit Rinderserumalbumin
(5%) in Tris-gepufferter Kochsalzlösung mit Tween-20 (0,05%) (TBST)
blockiert. Die Blots wurden mit einem monoklonalen Antikörper (1
: 1.000 Verdünnung)
gegen Phosphotyrosin (Upstate Biotechnology) für mindestens 1 Stunde bei Raumtemperatur
getestet. Die Blots wurden mit TBST gewaschen, mit einem Antikörper gegen
Mausimmunoglubolin G, konjugiert gegen alkalische Phosphatase (Promega) (verdünnt 1 :
7.500) für
mindestens 30 Minuten bei Raumtemperatur überprüft. Reaktive Banden wurden
visualisiert mit 5-Brom-4-chlor-3-indoyl-1-phosphat und Nitro-Blautetrazolium.
Immunblots wurden mit einem Scan Jet Plus (Hewlett-Packard) Densitometer
gescannt. Die Intensitäten
der Signale für
die nicht-stimulierten MCF-7 Zellen betrugen 20 bis 30 Einheiten.
Voll stimulierte, von p185erbB2 abstammende
Signale hatten 180 bis 200 Einheiten. Der 0,6 M NaCl Pool, welcher
die meiste Aktivität
enthielt, wurde auf einer Polyaspartinsäure (PolyLC)-Säule aufgetragen,
die mit 17 mM Natriumphosphat (pH 6,8), welches Ethanol (30%) enthielt, äquilibriert
war. Ein linearer Gradient von 0,3 M bis 0,6 M NaCl in dem Äquilibrierungspuffer
wurde verwendet, um die gebundenen Proteine zu eluieren. Ein Aktivitätspeak (bei
ca. 0,45 M NaCl) wurde ferner auf einer C4-Umkehrsäule (SynChropak
RP-4) weiter fraktioniert,
die mit Puffer, der TFA (0,1%) und Acetonitril (15%) enthielt, äquilibriert
war. Die Proteine wurden von dieser Säule mit einem Acetonitrilgradienten
von 25 bis 40% über
60 Minuten eluiert. Die Fraktionen (1 ml) wurden gesammelt, hinsichtlich
ihrer Aktivität
getestet und mit SDS-PAGE
auf Tris-Glycingelen (4–20%,
Novex) analysiert.
-
HPLC
gereinigte HRG-α wurde
mit Lysin C in SDS (0,1%), 10 mM Dithiothreitol, 0,1 M NH4HCO3 (pH 8,0) für 20 Stunden
bei 37°C
verdaut und die erzielten Fragmente wurden auf einer Synchrom C4-Säule aufgelöst (4.000
A, 0,2–10
cm). Die Säule
war in 0,1% TFA äquilibriert
und wurde mit einem 1-Propanolgradienten in 0,1% TFA (W. J. Henzel,
J. T. Stults, C. Hsu, D. W. Aswad, J. Biol. Chem. 264, 15905 (1989))
eluiert. Die Peaks von dem chromatographischen Lauf wurden unter
Vakuum getrocknet und sequenziert. Eines der Peptide (eluierend
bei etwa 24% 1-Propanol) gab die Sequenz [A]AEKEKTF[C]VNGGEXFMVKDLXNP
(SEQ ID Nr. 162). Die Reste in den Klammern waren nicht sicher und
ein X stellt einen Zyklus dar, bei dem es nicht möglich war,
die Aminosäure
zu identifizieren. Der Anfangsertrag betrug 8,5 pMol und die Sequenz
entsprach keinem bekannten Protein. Die Reste 1, 9, 15 und 22 wurden
später
als Cystein in der cDNA-Sequenz identifiziert. Eine direkte Sequenzierung
der etwa 45 kD Bande von dem Gel, welches überladen war und auf eine PVDF
Membran geblottet wurde, ergab eine geringe Häufigkeitssequenz XEXKE[G][R]GK[G]K[G]KKKEXGXG[K]
(SEQ ID Nr. 30) mit einem sehr geringen anfänglichen Ertrag (0,2 pMol).
Diese entsprach den Aminosäureresten
2 bis 22 von Heregulin-α (31), was vermuten lässt, dass Serin 2 der NH2-Terminus von proHRG-α ist. Obgleich der NH2-Terminus blockiert war, wurde beobachtet,
dass gelegentlich eine geringe Menge eines normal blockierten Proteins
nicht post-translational modifiziert werden kann.
-
Die
NH2 terminate Stellung wurde durch Massenspektrometrie
des Proteins nach Verdau mit Cyanogenbromid bestätigt. Der COOH-Terminus des
isolierten Proteins wurde nicht definitiv identifiziert. Durch Mischungssequenzierung
der proteolytischen Fragmente scheint jedoch die Reifesequenz sich
nicht über
den Rest 241 hinaus zu erstrecken. Die Abkürzungen für die Aminoreste sind: A, Ala;
C, Cys; D, Asp; E, Glu; F, Phe; G, Gly; H, His; I, Ile; K, Lys;
L, Leu; M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; 5, Ser; T, Thr;
V, Val; W, Trp; und Y, Tyr.
-
Als
eine Quelle der cDNA-Klone wurde eine Oligo(dT)-primed λgt10 (T.
V. Huynn, R. A. Young, R. W. Davis, λgt10 und λgt11 DNA Cloning Techniques:
A Practical Approach, D. Glover, Ed. (ICR Press, Oxford, (1984))
cDNA-Bibliothek hergestellt (U. Gubler und B. J. Hoffman, Gene 25,
263 (1983)) mit gereinigter mRNA (J. M. Chirwin, A. E. Przbyla,
R. J. MacDonald, W. J. Rutter, Biochemistry 18, 5294 (1979)) aus
MDA-MB-231 Zellen. Das folgende, achtfach degenerierte Antisens-Deoxyoligonukleotid,
welches die 13-Aminosäuresequenz
AEKEKTFCVNGGE (SEQ ID Nr. 31) (13) codiert, wurde aufgebaut auf
Basis der menschlichen Codonhäufigkeitsoptima
(R. Lathe, J. Mol. Biol. 183, 1 (1985)) und chemisch synthetisiert:
5'-CTCGCC (G oder
T) CC (A oder G) TTCAC (A oder G) CAGAAGGTCTTCTCCTTCTCAGC-3' (SEQ ID Nr. 40).
Für den
Zweck eines Sondenaufbaus wurde ein Cystein für einen unbekannten Rest in
der Aminosäuresequenz
verwendet. Die Sonde wurde durch Phosphorilierung markiert und unter
Niedrigstringensbedingungen mit der cDNA-Bibliothek hybridisiert.
Das proHRG-α Protein
wurde in dieser Bibliothek identifiziert. HRB-β1 cDNA wurde durch Testung einer
sekundären
Oligo(dT)-primed λgt10-Bibliothek
identifiziert, die aus MDA-MB-231Zell-mRNA hergestellt wurde, mit
Sequenzen, die von den 5' und
3' Enden von proHRG-α abstammten.
Der Klon 13 (2A) war ein Produkt der Testung
eines primed (5'-CCTCGCTCCTTCTTCTTGCCCTTC-3' Primers (SEQ ID
Nr. 41); proHRG-α Antisens-Nukleotide
33 bis 56) MDA-MB-231 λgt10
Bibliothek mit 5' HRG-α Sequenz. Eine
Sequenz, die dem 5'-Ende
des Klons 13 entsprach, wurde als eine Sonde verwendet für die Identifizierung
von proHRGβ2
und proHRGβ3
in einer dritten Oligo(dT)-primed λgt10 Bibliothek, die von MDA-MB-231 Zell-mRNA abstammte. Zwei
cDNA Klone, die für
jede der vier HRGs codieren, wurden sequenziert (F. Sanger, S. Milken,
A. R. Coulson, Proc. Natl. Acad. Sci. U.S.A. 74, 5463 (1977)). Eine
andere, als Klon 84 bezeichnete cDNA hatte eine Aminosäuresequenz,
die bis zur Aminosäure
420 mit proHRGβ2
identisch war. Dem Stoppcodon in der Position 421 folgt eine unterschiedliche
3' nicht-translatierte Sequenz.
-
Beispiel 9
-
Isolierung
einer weiteren Spleißvariante
-
Die
Verfahren nach Beispiel 6 ergaben vier nahe verwandte Sequenzen
(Heregulin α, β1, β2, β3), die als
ein Ergebnis der Spleißvariation
entstanden sind. (Peles et al. (Cell 69, 205 (1992) und Wen et al.
(Cell 69, 559 (1992)) haben eine andere Spleißvariante (von der Ratte) unter
Verwendung eines Reinigungs- und Klonierungsansatzes isoliert, der ähnlich zu
dem ist, der in den Beispielen 1 bis 4 und 6 beschrieben ist, wobei
ein Protein, dass an p185erbB2 bindet, beteiligt
ist. Der cDNA-Klon wurde wie folgt erzielt (über die Reinigung und Sequenzierung
eines p185erbB2 Bindungsproteins von einer
transformierten Fibroplastenzelllinie der Ratte). Ein p185erbB2 Bindungsprotein wurde aus konditioniertem
Medium wie folgt gereinigt. Die konditionierten Medien von drei
Ernten von 500 Rollerflaschen (insgesamt 120 Liter) wurden gesammelt
und durch Filtration durch 0,2 μ Filter
gereinigt und 31-fach mit einem Pelicon-Ultrafiltrationssystem unter
Verwendung von Membranen mit einer molekularen Größenabsperrung
von 20 kD konzentriert. Alte Reinigungsschritte wurden durchgeführt unter
Verwendung eines schnellen-Protein-Flüssigkeitschromatographiesystems
von Pharmacia. Das konzentrierte Material wurde direkt auf eine
Säule von
Heparin-Celtulose (150 ml, preäquilibriert
mit phosphatgepufferter Kochsalzlösung (PBS)) gegeben. Die Säule wurde
mit PBS, das 0,2 M NaCl enthielt, gewaschen, bis keine Extinktion
bei 280 nm Wellenlänge
entdeckt werden konnte. Die gewonnenen Proteine wurden dann mit einem
kontinuierlichen Gradienten (250 ml) von NaCl (von 0,2 M bis 1,0
M) eluiert und 5 ml Fraktionen wurden gesammelt. Proben (0,01 ml)
der gesammelten Fraktionen wurden verwendet für den quantitativen Test der Kinase
stimulierenden Aktivität.
Aktive Fraktionen von drei Säulenläufen (Gesamtvolumen
360 ml) wurden zusammengegeben, auf 25 ml unter Verwendung einer
YM10-Ultrafiltrationsmembran (Amicon, Danvers, MA) konzentriert
und Ammoniumsulfat wurde zugegeben, um eine Konzentration von 1,7
M zu erzielen. Nach Reinigung durch Zentrifugation (10.000 × g, 15
Minuten) wurde das gepoolte Material auf eine Phenyl-Superose-Säule geladen
(HR10/10, Pharmacia). Die Säule
wurde entwickelt mit einem 45 ml Gradienten aus (NH4)2SO4 (von 1,7 M bis
kein Salz) in 0,1 M Na2PO4 (pH
7,4) und 2 ml Fraktionen wurden gesammelt und getestet (0,002 ml
pro Probe) für
die Kinasestimulierung (wie im Beispiel 6 beschrieben). Der Hauptpeak
der Aktivität
wurde zusammengefasst und gegen 50 mM Natriumphosphatpuffer (pH
7,3) dialysiert. Eine Mono-S-Kationenaustauschersäule (HR5/5,
Pharmacia) wurde preäquilibriert
mit 50 mM Natriumphosphat. Nach Beladung des aktiven Materials (0,884
mg Protein; 35 ml) wurde die Säule
mit dem Startpuffer gewaschen und dann entwickelt mit einer Rate
von 1 ml/Minute mit einem Gradienten von NaCl. Die stimulierende
Kinaseaktivität
wurde bei 0,45–0,55
M Salz entdeckt und über
vier Fraktionen von jeweils 2 ml verteilt. Diese wurden zusammengefasst
und direkt auf eine Cu+2 Chelatsäule (1,6
ml, HR2/5 Chelatsuperose, Pharmacia) geladen. Der Großteil des
Proteins adsorbierte an dem Harz und wurde graduell eluiert mit
einem 30 ml linearen Gradienten aus Ammoniumchlorid (0–1 M). Die
Aktivität
eluierte in einem einzelnen Proteinpeak im Bereich von 0,05 bis 0,2
M NH4Cl. Proben von den verschiedenen Schritten
der Reinigung wurden mit Gelelektrophorese analysiert und anschließend mit
Silber gefärbt
unter Verwendung eines Kits von ICN (Costa Mesa, CA). Die Proteingehalte
wurden bestimmt mit einem Coomassie-Blau-Färbungstest unter Verwendung
eines Kits von Bio-Rad (Richmond, CA).
-
Das
p44-Protein (10 μg)
wurde in 200 μl
von 0,1 M Ammoniumbicarbonatpuffer (pH 7,8) wieder gelöst. Der
Verdau wurde durchgeführt
mit L-1-Tosyl-Amid-2-phenylethylchlormethylketon behandeltem Trypsin
(Serva) bei 37°C
für 18
Stunden bei einem Enzym-Substrat-Verhältnis von 1 : 10. Die erzielte
Peptidmischung wurde mit Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen getrennt und bei 215 nm unter Verwendung
einer Vydac C4 Mikrosäule
(2,1 mm Innendurchmesser × 15
cm, 300 A) und einem HP 1090 Flüssigkeitschromatographiesystems,
ausgerüstet
mit einem Diodenarraydetektor und einer Arbeitsstation, überwacht.
Die Säule
wurde mit 0,1% Trifluoressigsäure
(mobile Phase A) äquilibriert
und die Elution wurde mit einem linearen Gradienten von 0%–55% mobile
Phase B (90% Acetonitril in 0,1% Trifluoressigsäure) über 70 Minuten durchgeführt. Die
Fließrate
betrug 0,2 ml/Minute und die Säulentemperatur
betrug 25°C.
Ein dritter Aliquot der Peptidpeaks, die manuell von dem HPLC-System gesammelt
wurden, wurde charakterisiert durch N-terminate Sequenzanalyse durch
Edman-Abbau. Die nach 27,7 Minuten (T 27,7) eluierte Fraktion enthielt die
gemischten Aminosäuresequenzen
und wurde nach Reduktion wie folgt weiter chromatographiert. Ein
70% Aliquot der Peptidfraktion wurde im Vakuum getrocknet und in
100 μl von
0,2 M Ammoniumbicarbonatpuffer (pH 7,8) wieder gelöst. DTT
(Endkonzentration 2 mM) wurde der Lösung zugegeben, die dann für 30 Minuten bei
37°C inkubiert
wurde. Die reduzierte Peptidmischung wurde dann mit Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen unter Verwendung einer Vydac-Säule (2,1
mm Innendurchmesser × 15
cm) getrennt. Die Elutionsbedingungen und die Fließrate waren
identisch mit dem oben beschriebenen Verfahren. Die Analyse der
Aminosäuresequenz
des Peptides wurde durchgeführt
mit einem Modell 477 Proteinsequenziergerät (Applied Biosystems, Inc.,
Foster City, CA), das mit einem on-line Phenylthiohydantoin (PTH)-Aminosäureanalysator
und einem Modell 900 Datenanalysesystem ausgerüstet war (Hunkapiller et al.
(1986) In Methods of Protein Microchracterization, J. E. Shively,
ed. (Clifton, New Jersey: Humana Press; S. 223–247). Das Protein wurde auf
eine Glasfaserscheibe geladen, die mit Trifluoressigsäure behandelt
war und präcyclesiert
mit Polypren und NaCl. Die PTH-Aminosäureanalyse wurde mit einem
Mikroflüssigkeitschromatographiesystem (Modell
120) unter Verwendung von zwei Spritzpumpen und englumigen Umkehrphasen
(C-18)-Säulen
(Applied Biosystems, 2,1 mm × 250
mm) durchgeführt.
-
RNA
wurde isoliert aus Rat1-EJ Zellen durch Standardverfahren (Maniatis
et al., Molecular Cloning: A Laboratory Manual (Cold Spring Harbor,
New York (1982) und Poly (A)+ wurde selektiert
unter Verwendung eines mRNA-Separatorkits
(Clontech Lab, Inc., Palo Alto, CA). cDNA wurde synthetisiert mit
dem Superscript Kit (von BRL Life Technologies, Inc., Bethesda,
MD). Säulen-fraktionierte,
doppelsträngige
cDNA wurde in einen Sal1- und Not1-geschnittenen pJT-2 Plasmidvektor legiert,
ein Derivat von dem pCD-X Vektor (Okayama und Berg, Mol. Cell Biol.
3: 280 (1983), und transformiert in DH10B E. Coli Zellen durch Elektroporation
(Dower et al., Nucl. Acids Res. 16: 6127 (1988)). Etwa 5 × 105 primäre
Transformanten wurden mit zwei Oligonukleotidproben getestet, die
abstammten von den Proteinsequenzen des N-Terminus von NDF (Reste
5–24)
und von dem T40.4 tryptischen Peptid (Reste 7–12). Ihre entsprechenden Sequenzen
waren wie folgt (N zeigt alle 4 nt):
-
-
Die
synthetischen Oligonukleotide wurden endmarkiert mit (γ-32p)ATP mit T4 Polynukleotidkinase und wurden
verwendet, um Replikasets von Nitrocellulosefilter zu screenen.
Die Hybridisierungslösung
enthielt 6 × SSC,
50 mM Natriumphosphat (pH 6,8), 0,1% Natriumpyrophosphat, 2 × Denhardt's Lösung, 50 μg/ml Lachssperma-DNA
und 20% Formamid (für
Probe 1) oder kein Formamid (für
Probe 2). Die Filter wurden entweder bei 50°C mit 0,5 × SSC, 0,2% SDS, 2 mM EDTA
(für Probe
1) oder bei 37°C
mit 2 × SSC,
0,2% SDS, 2 mM EDTA (für
Probe 2) gewaschen. Die Autoradiographie der Filter ergab zehn Klone,
die mit beiden Sonden hybridisierten. Diese Klone wurden durch Wiederplattieren
und Probenhybridisierung, wie oben beschrieben, gereinigt.
-
Die
cDNA-Klone wurden sequenziert unter Verwendung eines Applied Biosystems
373A automatischer DNA Sequenzierer und Applied Biosystems Taq DyeDeoxyTM Terminator cycle sequencing Kits gemäß den Anweisungen
des Herstellers. In einigen Fällen
wurden die Sequenzen erzielt unter Verwendung von [35S]dATP
(Amersham) und SequenaseTM Kits von U.S.
Biochemicals gemäß den Anweisungen
des Herstellers. Beide Stränge
des cDNA-Klons 44 wurden verwendet unter Verwendung der synthetischer
Oligonukleotide als Primer. Die Sequenz der meisten 5' 350 nt wurde in
sieben unabhängigen
cDNA-Klonen bestimmt. Der erzielte Klon zeigte das Muster, welches
in der 30 (NDF) gezeigt ist.
-
Beispiel 10
-
Strategien
für den
Nachweis von anderen möglichen
Spleißvarianten
-
Die
Ausrichtung der abgeleiteten Aminosäuresequenzen von den cDNA-Klonen und den PCR-Produkten
vom Rind mit den veröffentlichten
menschlichen (31) und Rattensequenzen
zeigen ein hohes Maß an Ähnlichkeit,
was anzeigt, dass diese Sequenzen von homologen Genen innerhalb
der drei Arten abstammen. Die variable Zahl der Boten-RNA-Transkripte, die
auf dem cDNA/PCR Produktenniveau nachweisbar ist, beruht wahrscheinlich
auf einem intensiven gewebespezifischen Splicing. Die erzielten
und in der 30 gezeigten Muster lassen
vermuten, dass andere Spleißvarianten
existieren. Eine Liste von wahrscheinlichen Spleißvarianten
ist in der 37 angezeigt. Viele von
diesen Varianten können
durch spezifische codierende Segmenttestung von cDNA-Bibliotheken,
die von verschiedenen Geweben abstammen, und durch PCR-Experimente
unter Verwendung von Primerpaaren, die spezifisch zu besonderen
codierenden Segmenten sind, erzielt werden. Alternativ können die
Varianten von spezifischen cDNA-Klonen,
PCR-Produkten oder genomischen DNA-Regionen über einen Fachmann bekannte
Schneid- und Spleißtechniken
zusammengestellt werden. Zum Beispiel kann eine seltene Stelle zum
Schneiden mit einem Restriktionsenzym in einem allgemein codierenden
Segment (zum Beispiel A) verwendet werden, um den FBA-Aminoterminus
von GGF2BPP5 mit den terminalen Carboxysequenzen von GGF2BPP1, GGFBPP2,
GGFBPP3 oder GGFBPP4 zu verbinden. Wenn die Gegenwart oder die Abwesenheit
von den codierenden Segmenten E und/oder G einen Vorteil für die betrachteten
und angegebenen Verwendungen bereitstellt, können diese codierenden Segmente
in die Expressionskonstrukte aufgenommen werden. Diese Variantensequenzen
können
in rekombinanten Systemen exprimiert werden und die rekombinanten
Produkte können
getestet werden, um die mitogenetische Aktivität auf Schwann-Zellen zu bestimmen
sowie hinsichtlich der Fähigkeit
an den p185erbB2 Rezeptor zu binden und diesen
zu aktivieren.
-
Beispiel 11
-
Identifikation von funktionalen
Elementen von GGF
-
Die
abgeleiteten Strukturen der Familie der GGF-Sequenzen zeigen, dass
die längsten
Formen (repräsentiert
durch GGF2BPP4) für
Transmembranproteine codieren, wo der extrazelluläre Teil
eine Domäne enthält, die
dem epidermalen Wachstumsfaktor ähnelt
(siehe Carpenter und Wahl in Peptide Growth Factors and Their Receptors
I, Seite 69–133,
Springer-Verlag, NY 1991). Die Positionen der Cysteinreste in den
codierenden Segmenten C und C/D oder C/D' Peptidsequenzen sind konserviert bezüglich den
analogen Resten in der Peptidsequenz des epidermalen Wachstumsfaktors
(EGF) (siehe 35, SEQ ID Nrn. 151–153). Dies lässt vermuten,
dass die extrazelluläre
Domäne
als Rezeptorerkennungs- und biologische Aktivierungsstelle fungiert.
Verschiedene Variantenformen fehlen, wie H, K und L codierende Segmente
und können
somit als sekreierte, diffusionsfähige, biologisch aktive Proteine
exprimiert werden. Die GGF DNA-Sequenzen, die für Polypeptide codieren, welche
die EGF-änliche
Domäne
(EGFL) umfassen, können
eine volle biologische Aktivität
für die
Stimulierung der mitogenetischen Glialzellaktivität besitzen.
-
Membrangebundene
Versionen von diesem Protein können
die Schwann-Zellproliferation
induzieren, wenn sie auf der Oberfläche von Neuronen während der
Embryogenese oder während
der Nervenregeneration exprimiert werden (wo die Oberflächen der
Neuronen eng assoziiert sind mit den Oberflächen der proliferierenden Schwann-Zellen).
Sekregierte (nicht Membran gebundene) GGFs können als klassisch diffusionsfähige Faktoren
wirken, die mit den Schwann-Zellen in einiger Entfernung von ihrem
Sekretionsort interagieren können.
Andere Formen können
durch Gewebeverletzung und Zellzerstörung freigesetzt werden. Ein
Beispiel für
ein sekregiertes GGF ist das Protein, das durch GGF2HBS5 (siehe
Beispiel 6) codiert wird. Dies ist das einzig bekannte GGF, das
aus der Zelle (Beispiel 7) herausgeführt wird. Sekretion ist wahrscheinlich über eine N-terminate,
hydrophobe Sequenz gesteuert, die nur in der Region E gefunden wird,
was die N-terminale Domäne
darstellt, die innerhalb des rekombinanten GGF-II, das von GGF2HBS5
codiert wird, enthalten ist.
-
Andere
GGFs scheinen nicht sekregiert zu werden (siehe Beispiel 6). Diese
GGFs können
Formen auf Verletzungsreaktionen sein, die als Konsequenz einer
Gewebeschädigung
freigesetzt werden.
-
Andere
Regionen der prognostizierten Proteinstruktur von GGF-II (codiert
von GGF2HBS5) und andere Proteine, welche die Region B und A enthalten,
zeigen Ähnlichkeiten
zu dem menschlichen Basalmembran-Heparan-Sulfat-Proteoglycan-Kernprotein (Referenz).
Das Peptid ADSGEY, das als nächstes
zu dem zweiten Cystein der C2 Immunoglobulinfalte in diesen GGFs
lokalisiert ist, tritt in neun von zweiundzwanzig C-2 Wiederholungen
auf, die in diesem Basalmembranprotein gefunden werden. Dieser Hinweis
lässt stark vermuten,
dass diese Proteine mit Matrixproteinen assoziiert sein können, wie
solche, die mit Neuronen und der Glial assoziiert sind und es kann
ein Verfahren für
die Absonderung von Gliawachstumsfaktoren an den Zielorten vermutet
werden.
-
Beispiel 12
-
Reinigung von GGFs aus
rekombinanten Zellen
-
Um
die volle Länge
oder Teile von GGFs zu erzielen für den Test auf biologische
Aktivität,
können
die Proteine unter Verwendung von klonierter DNA überproduziert
werden. Verschiedene Ansätze
können
angewandt werden. Eine rekombinante E. Coli Zelle, welche die oben
beschriebenen Sequenzen enthält,
kann hergestellt werden. Expressionssysteme, wie pNH8a oder pHH16a
(Stratagene, Inc.), können
für diesen
Zweck gemäß den Anweisungen
des Herstellers verwendet werden. Alternativ können diese Sequenzen in einen Säugetierexpressionsvektor
eingefügt
werden und eine überproduzierende
Zelllinie kann so aufgebaut werden. Für diesen Zweck wurde zum Beispiel
DNA, die für
einen GGF codiert, Klon GGF2BPP5, in COS-Zellen und in Chinesischen Hamstereizellen
exprimiert (siehe Beispiel 7) (J. Biol. Chem. 263, 3521–3527, (1981)). Dieser
Vektor enthält
die GGF DNA-Sequenzen
und kann in Wirtszellen unter Verwendung von etablierten Verfahren
transfiziert werden.
-
Die
transiente Expression kann überprüft werden
oder G418 resistente Klone können
in Gegenwart von Methotrexat wachsen, um solche Zellen auszuwählen, die
das dhfr Gen amplifizieren (enthaltend auf dem pMSXND Vektor) und
bei dem Verfahren die benachbarte GGF Protein codierende Sequenz
co-amplifizieren. Da die CHO-Zellen in einem total serumfreien,
proteinfreien Medium (Hamilton und Ham, in vitro 13, 537–547 (1977)) gehalten
werden können,
kann das gewünschte
Protein aus dem Medium gereinigt werden. Western-Analyse unter Verwendung
der im Beispiel 9 hergestellten Antiseren kann verwendet werden,
um die Gegenwart des gewünschten
Proteins in dem konditionierten Medium der überproduzierenden Zellen nachzuweisen.
-
Das
gewünschte
Protein (rGGF-II) wurde aus dem Medium gereinigt, das durch transientexprimierende
COS-Zellen konditioniert war. rGGF-II wurde von dem konditionierten
Medium geerntet und partiell gereinigt unter Verwendung der Kationenaustauscherchromatographie
(POROS-HS). Die Säule
wurde äquilibriert
mit 33,3 mM MES, pH 6,0. Konditionierte Medien wurden mit einer
Fließrate
von 10 ml/Minute geladen. Der Peak, der die Schwann-Zellproliferationsaktivität enthielt
und immunreaktiv war (unter Verwendung der polyklonalen Antiseren
gegen ein oben beschriebenes GGFII Peptid) wurde eluiert mit 50
mM Tris, 1 M NaCl, pH 8,0, (50A und 50B).
-
rGGF-II
wurde auch unter Verwendung einer stabilen Eizelllinie vom Chinesischen
Hamster exprimiert. rGGF-II von den geernteten, konditionierten
Medien wurde teilweise gereinigt unter Verwendung der Kationenaustauscherchromatographie
(POROS-HS). Die Säule
wurde äquilibriert
mit PBS, pH 7,4. Die konditionierten Medien wurden bei 10 ml/Minute
beladen. Der Peak, der die Schwann-Zellproliferationsaktivität und die
Immunoreaktivität
enthielt (unter Verwendung der GGFII polyklonalen Antiseren) wurde
eluiert mit 50 mM Hepes, 500 mM NaCl, pH 8,0. Ein zusätzlicher
Peak wurde beobachtet bei 50 mM Hepes, 1 M Na Cl, pH 8,0, mit sowohl
Proliferation als auch Immunoreaktivitätsaktivität (51).
-
rGGF-II
kann ferner gereinigt werden unter Verwendung der hydrophoben Interaktionschromatographie
als ein hochauflösender
Schritt, der Kationenaustauscher/Umkehrphasenchromatographie (falls
notwendig als zweiter hochauflösender
Schritt), eines viralen Inaktivitätsschrittes und eines DNA-Entfernungsschrittes,
wie eine Anionenaustauscherchromatographie.
-
Es
folgt eine detaillierte Beschreibung der verwendeten Verfahren:
Die
Schwann-Zellproliferationsaktivität des rekombinanten GGF-II
Peaks, der von der Kationenaustauschersäule eluiert wurde, wurde wie
folgt bestimmt: Die mitogenetischen Reaktionen der kultivierten
Schwann-Zellen wurden gemessen in Gegenwart von 5 M Forskolin unter
Verwendung des Peaks, der bei 50 mM Tris, 1 M NaCl, pH 8,0, eluiert
wurde. Der Peak wurde zugegeben zu 20 1, 10 1 (1 : 10) 10 1 und
(1 : 100) 10 1. Die Einlagerung von 125I-Uridin
wurde bestimmt und nach 18 bis 24-stündiger Exposition exprimiert
als (CPM).
-
Ein
Immunoblot unter Verwendung eines polyklonalen Antikörpers, der
gegen ein Peptid von GGF-II gebildet wurde, wurde wie folgt durchgeführt: 10 μl von verschiedenen
Fraktionen liefen auf 4–12
Gradientengelen. Die Gele wurden auf Nitrocellulosepapier übertragen
und die Nitrocelluloseblots wurden mit 5% BSA blockiert und gegen
GGF-II spezifischen Antikörper
(1 250 Verdünnung)
entwickelt. 125I Protein A (1 : 500 Verdünnung, spezifische
Aktivität
= 9,0/Ci/g) wurde als zweiter Antikörper verwendet. Die Immunoblots
wurden auf Kodak Röntenfilmen
für 6 Stunden
entwickelt. Die Peakfraktionen, die mit 1 M NaCl eluierten, zeigten
ein breites immunoreaktives Band bei 65–90 kD, was der erwartete Größenbereich
für die
GGFII und die Glycoformen mit höheren
Molekulargewichten ist.
-
Die
GGF-II Reinigung auf den Kationenaustauschersäulen wurde wie folgt durchgeführt: Konditionierte
Medien von CHO-Zellen, welche rGGFII exprimierten, wurden auf die
Kationenaustauschersäule
mit 10 ml/Minute geladen und die Säule wurde äquilibriert mit PBS, pH 7,4.
Die Elution wurde erzielt mit 50 mM Hepes, 500 mM NaCl, pH 8,0,
und 50 mM Hepes, 1 M NaCl, pH 8,0. Alle Fraktionen wurden unter
Verwendung des hier beschriebenen Schwann-Zellproliferationstests
(CPM) analysiert. Die Proteinkonzentration (mg/ml) wurde unter Verwendung
des Bradford Tests unter Verwendung von BSA als Standard bestimmt.
-
Ein
Western-Blot unter Verwendung von 10 μl von jeder Fraktion wurde durchgeführt. Wie
in den 51A und 51B angezeigt,
comigrierte die Immunoreaktivität
und die Schwann-Zellaktivität.
-
Der
hier beschriebene, mitogenetische Schwann-Zelltest kann verwendet
werden, um das exprimierte Produkt des Klons mit voller Länge oder
jeden biologisch aktiven Teil davon zu testen. Der Klon mit voller
Länge GGF2BPP5
wurde transient in COS-Zellen exprimiert. Intrazelluläre Extrakte
von transfizierten COS-Zellen zeigen eine biologische Aktivität, wenn
sie in dem in Beispiel 1 beschriebenen Schwann-Zellproliferationstest überprüft wurden.
Ferner wurde der GGF2HBS5 codierende Klon mit voller Länge transient
in CHO und in Insekten (Beispiel 7)-Zellen exprimiert. In diesem
Fall zeigten sowohl der Zellextrakt als auch die konditionierten Medien
eine biologische Aktivität
in dem in Beispiel 1 beschriebenen Schwann-Zellproliferationstest.
Jedes Mitglied der spleißvarianten
Familie von komplementärer
DNAs, die von dem GGF-Gen (einschließlich den Heregulinen), kann
auf diese Weise exprimiert und in dem Schwann-Zellproliferationstest
durch einen Fachmann getestet werden.
-
Alternativ
kann das rekombinante Material von anderen Varianten gemäß Wen et
al. (Cell 69, 559 (1992)) isoliert werden, die die Splicingvariante
neu Differenzierungsfaktor (NDF) in COS-7 Zellen exprimierten. CDNA
Klone, die in dem pJT-2 eukaryontischen Plasmidvektor inseriert
sind, sind unter der Kontrolle des frühen SV40 Promotors und werden
3'-flankiert von
den SV40 Terminierungs- und Polyadenylierungssignalen. COS-7 Zellen
wurden mit der pJT-2 Plasmid-DNA durch Elektroporation wie folgt
transfeziert: 6 × 106 Zellen (in 0,8 ml DMEM und 10% FEBS) wurden
in eine 0,4 cm Cuvette überführt und
gemischt mit 20 μg
Plasmid-DNA in 10 μl
TE Lösung
(10 mM Tris-HCl (pH 8,0), 1 mM EDTA). Die Elektroporation wurde
bei Raumtemperatur und bei 1.600 V und 25 μF unter Verwendung eines Bio-Rad
Gene Pulser Gerätes
durchgeführt,
wobei die Pulskontrolleinheit auf 200 Ohm eingestellt wurde. Die
Zellen wurden dann in 20 ml DMEM, 10% FBS verdünnt und in eine T75 Flasche
(Falcon) überführt. Nach
14 Stunden Inkubation bei 37°C
wurde das Medium ersetzt durch DMEM, 1% FBS und die Inkubation wurde
für weitere
48 Stunden fortgeführt.
Konditioniertes Medium, das rekombinantes Protein enthielt, wurde
von den Zellen geerntet, was die biologische Aktivität in einer Zelllinie
anzeigt, die den Rezeptor für
dieses Protein exprimiert. Diese Zelllinie (kultivierte, menschliche
Brustkarzinomzelllinie AU 565) wurde mit dem rekombinanten Material
behandelt. Die behandelten Zellen zeigten eine morphologische Veränderung,
was charakteristisch für
die Aktivierung des erbB2 Rezeptors ist. Konditioniertes Medium
von diesem Typ kann auch in dem Schwann-Zellproliferationstest getestet
werden.
-
Beispiel 13 Reinigung
und Test der anderen Proteine, die den p185erbB2 Rezeptor
binden
-
I. Reinigung von gp30
und p70
-
Lupu
et al. (Science 249, 1552 (1990)) und Lippman und Lupu (Patentanmeldung
Nr. PCT/US91/03443 (1990)), hiermit durch Verweis eingeführt, haben
ein Protein aus konditioniertem Medium einer menschlichen Brustkrebszelllinie,
MDA-MB-231, wie folgt gereinigt.
-
Das
Sammeln der konditionierten Medien wurde unter Verwendung von bekannten
Verfahren durchgeführt.
Die Medien wurden 100-fach in einer Amicon Ultrafiltrationszelle
(YM5 Membran) (Amicon, Danvers, MA) konzentriert. Nach Reinigung
und Konzentrierung wurden die Medien bei –20°C gelagert, wobei aufeinander
folgende Sammlungen während
den folgenden Tagen gemacht wurden. Die konzentrierten Medien wurden
dialysiert unter Verwendung von Spectra/por® 3
Tubing (Spectrum Medical Industries, Los Angeles, CA) gegen 100
Volumen von 0,1 M Essigsäure über einen
zwei Tageszeitraum bei 4°C.
Das Material, welches während
der Dialyse gefällt
wurde, wurde durch Zentrifugation bei 4.000 Umdrehungen pro Minute
für 30
Minuten und bei 4°C
entfernt. Proteaseinhibitoren wurden zugesetzt. Die gereinigte Probe
wurde dann lyophilisiert.
-
Lyophilisiertes,
konditioniertes Medium wurde in 1 M Essigsäure bis zu einer Endkonzentration
von etwa 25 mg/ml Gesamtprotein gelöst. Unlösliches Material wurde durch
Zentrifugation bei 10.000 Umdrehungen pro Minute für 15 Minuten
entfernt. Die Probe wurde dann auf eine Sephadex G-100 Säule (XK
16, Pharmacia, Piscataway, NJ) geladen, äquilibriert und mit 1 M Essigsäure bei
4°C mit
einem Aufwärtsstrom
von 30 ml/Stunde eluiert. 100 ng Protein wurde aus 4 ml von 100-fach
konzentriertem Medium gewonnen. Fraktionen, die 3 ml des Eluates
enthielten, wurden lyophilisiert und resuspendiert in 300 μl PBS für den Test
und dienten als Quelle für
die weitere Reinigung.
-
Sephadex
G-100 gereinigtes Material lief auf einer Hochdruck-Flüssigkeitschromatographie
mit umgekehrten Phasen (HPLC). Der erste Schritt umfasste einen
steilen Acetonitrilgradienten. Der steile Acetonitrilgradient und
alle anderen HPLC-Schritte wurden bei Raumtemperatur nach Äquilibrierung
der C3-Umkehrphasensäule
mit 0,05% TFA (Trifluoressigsäure)
in Wasser (HPLC-Qualität)
durchgeführt.
Die Proben wurden geladen und die Fraktionen wurden mit einem linearen
Gradienten (0–45%
Acetonitril in 0,05% TFA) bei einer Fließrate von 1 ml/Minute über einen
Zeitraum von 30 Minuten eluiert. Die Extinktion wurde bei 280 nm
aufgenommen. Ein ml Fraktionen wurden gesammelt und lyophilisiert
vor Analyse auf die EGF-Rezeptor-Konkurrenzreaktion.
-
Ein
zweiter HPLC-Schritt umfasste einen flachen Acetonitrilgradienten.
Der Pool der aktiven Fraktionen von dem vorherigen HPLC-Schritt
wurde über
die gleiche Säule
wieder chromatographiert. Die Elution wurde durchgeführt mit
einem 0–18%
Acetonitrilgradienten in 0,05% TFA über einen Zeitraum von 5 Minuten, worauf
sich ein linearer 18–45%
Acetonitrilgradient in 0,05% TFA über einen Zeitraum von 30 Minuten
anschloss. Die Fließrate
betrug 1,0 ml/Minute und 1 ml Fraktionen wurden gesammelt. Der menschliche
TGFα-ähnliche Faktor wurde bei einer
30–32%
Acetonitrilkonzentration als ein einzelner Peak, nachweisbar durch RRA,
eluiert.
-
Lupu
et al. (Proc. Natl. Acad. Sci. 89, 2287 (1992)) reinigten ein anderes
Protein, das an den p185erbB2 Rezeptor bindet.
Dieses besondere Protein, p75, wurde aus konditioniertem Medium
gereinigt, das für
das Wachstum von SKBr-3 (eine menschliche Brustkrebszelllinie) verwendet
wurde.
-
Die
Zelllinie wurde vermehrt in verbessertem Eagle's Medium (IMEM: GIBCO), ergänzt mit
10% fötalem
Rinderserum (GIBCO). Das Protein p75 wurde gereinigt aus konzentriertem
(100-fach), konditioniertem Medium unter Verwendung einer p185erbB2 Affinitätssäule. Die 94 Kilodalton extrazelluläre Domäne von p185erbB2 (die p75 bindet) wurde über eine
rekombinante Expression hergestellt und an eine Polyacrylamid-Hydrazido-Sepharose
Affinitätschromatographiematrix
gebunden. Nach Bindung wurde die Matrix intensiv mit eiskalter,
1,0 M HCl gewaschen und die Kügelchen
wurden mit 0,5 M NaNO2 aktiviert. Die Temperatur
wurde auf 0°C
für 20
Minuten gehalten. Anschließend
wurde filtriert und mit eiskalter 0,1 M HCl gewaschen. 500 ml an
konzentriertem, konditioniertem Medium lief aufgrund der Schwerkraft
durch die Kügelchen.
Die Säule
wurde gewaschen und stufenweise eluiert mit 1,0 M Zitronensäure bei
pH-Werten von 4,0 bis 2,0 (um die Dissozation von erbB2 und p75
zu erlauben). Alle Fraktionen wurden auf Pharmacia PD10 Säulen entsalzt.
Die Reinigung ergab ein homogenes Polypeptid von 75 kDa bei einem
3,0–3,5
Elutions-pH (bestätigt
durch Analyse auf SDS/PAGE durch Silberfärbung).
-
II. Bindung von gp30 an
p185erbB2
-
Das
gereinigte gp30 Protein wurde in einem Test überprüft, um zu bestimmen, ob es
an p185erbB2 bindet. Ein Konkurrenztest
mit einem monoklonalen Antikörper
gegen p185erbB2. Das gp30 Protein verdrängte die Antikörperbindung
an p185erbB2 in SK-BR-3 und MDA-MB-453 Zellen
(menschliche Brustkarzinomzelllinien, die den p185erbB2 Rezeptor
exprimieren). Die Schwann-Zellproliferationsaktivität von gp30
kann auch nachgewiesen werden durch Behandlung der Schwann-Zellkulturen
mit gereinigtem gp30 unter Verwendung des in den Beispielen 1–3 beschriebenen
Testverfahrens.
-
III. Bindung von p75 an
p185erbB2
-
Um
zu bestimmen, ob das 75 kDa Polypeptid (p75), das aus dem SKBr-3
konditioniertem Medium erzielt worden war, tatsächlich ein Ligand für das erbB2
Onkoprotein in den SKBr-3 Zellen ist, wurde ein Verdrängungstest,
wie oben für
gp30 beschrieben, durchgeführt.
-
Es
wurde gefunden, dass das p75 eine Bindungsaktivität zeigte,
wobei das Material von anderen Chromatographiefraktionen solch eine
Aktivität
nicht zeigte (Daten nicht gezeigt). Das Durchflussmaterial zeigte
einige Bindungsaktivität.
Dies kann aufgrund der Gegenwart von ausgeschüttetem erbB2 ECD erfolgt sein.
-
IV. Andere p185erbB2 Liganden
-
Peles
et al. (Cell 69, 205 (1992)) haben auch einen p185erbB2 stimulierenden
Liganden aus Rattenzellen gereinigt (NDF, siehe Beispiel 8 für die Methode).
Holmes et al. (Science 256, 1205 (1992)) haben Heregulin α aus menschlichen
Zellen gereinigt, das an 185erbB2 bindet
und dieses stimuliert (siehe Beispiel 6). Tarakovsky et al. Onkogene
6: 218 (1991) haben die Bindung eines 25 kD Polypeptides, das aus
aktivierten Makrophagen isoliert worden war, an den neu Rezeptor
gezeigt, eine p185erbB2 Homologie, hier
durch Verweis eingeführt.
-
VI. NDF-Isolierung
-
Yarden
und Peles (Biochemistry 30, 3543 (1991) haben ein 35 kD Glycoprotein
identifiziert, das den 185erbB2 Rezeptor
stimuliert. Das Protein wurde in konditioniertem Medium gemäß dem folgenden
Verfahren identifiziert. I-EJ Rattenzellen wuchsen bis zum Zusammenströmen in 175
cm2 Flaschen (Falcon). Einzelschichten wurden
mit PBS gewaschen und in einem serumfreien Medium für 10 bis
16 Stunden belassen. Das Medium wurde verworfen und durch frisches,
serumfreies Medium ersetzt, das nach dreitägiger Kultur gesammelt wurde.
Das konditionierte Medium wurde durch Zentrifugation bei geringer
Geschwindigkeit gereinigt und in einer Amicon-Ultrafiltrationszelle mit einer YM2-Membran
(Absperrung bei einem Molekulargewicht von 2.000) konzentriert.
Die biochemischen Analysen der neu stimulierenden Aktivität in dem
konditionierten Medium zeigten, dass der Ligand ein 35-kD Glycoprotein
ist, welches hitzestabil aber empfindlich gegenüber einer Reduktion ist. Der
Faktor ist entweder durch hohe Salzkonzentrationen oder durch sauren
Alkohol fällbar.
Eine partielle Reinigung des Moleküls durch selektive Fällung, Heparin-Agarose-Chromatographie und
Gelfiltration in verdünnter
Säure führte zu
einem aktiven Liganden, der zur Stimulierung des protoonkogenen
Rezeptors in der Lage ist, und der aber nicht wirksam ist auf tumorerzeugendes
neu Protein, welches konstitutiv aktiv ist. Die gereinigte Fraktion
zeigte jedoch die Fähigkeit
auch zur Stimulierung des verwandten Rezeptors von EGF, was vermuten
lässt,
dass diese zwei Rezeptoren durch einen bidirektionalen Mechanismus
funktional verbunden sind. Alternativ interagiert der angenommene
Ligand simultan mit beiden Rezeptoren. Die dargestellte biochemische
Charakterisierung des Faktors kann benutzt werden, um einen vollständig gereinigten
Faktor zu erzielen, mit dem diese Möglichkeiten zu testen sind.
-
In
anderen Publikationen beschreiben Davis et al. (Biochem. Biophys.
Res. Commun. 179, 1536 (1991), Proc. Natl. Acad. Sci. 88, 8582 (1991)
und Greene et al., PCT-Patentanmeldung PCT/US91/02331 (1990) die
Reinigung eines Proteins aus konditioniertem Medium einer menschlichen
T-Zell (ATL-2)-Zelllinie.
-
Die
ATL-2 Zelllinie ist eine IL-2 unabhängige HTLV-1 (+) T Zelllinie.
Mycoplasmenfreie ATL-2 Zellen wurden in RPMI 1640 Medium, das 10%
FCB enthielt, als Kulturmedium (10% FCS-RPMI 1640) bei 37°C in einer
befeuchteten Atmosphäre
mit 5% CO2 gehalten.
-
Für die Reinigung
der proteinartigen Substanz wurden die ATL-2 Zellen zweimal in 1 × PBS gewaschen
und zu 3 × 105 ml in serumfreiem RPMI 1640 Medium/2 mM
L-Glutamin kultiviert für
72 Stunden. Anschließend
wurden die Zellen pelletiert. Der Kulturüberstand, der so hergestellt
wurde, wird als „konditioniertes Medium" (C. M.) bezeichnet.
-
C.
M. wurde 100-fach von 1 Liter auf 10 ml konzentriert unter Verwendung
einer YM-2 Diaflo-Membran (Amicon, Boston, MA) mit einer Absprerrung
bei 1.000 Dalton. Zur Verwendung bei einigen Tests wurde konzentriertes
Kulturmedium, das Komponenten mit einem größeren Molekulargewicht als
1.000 enthielt, wieder auf das Ursprungsvolumen mit RPMI Medium
verdünnt.
Eine Gelelektrophorese unter Verwendung eines Polyacrylamid-Gradientengels
(Integrated Separation Systems, Hyde Park, MD oder Phorecast System
by Amersham, Arlington Heights, IL) mit anschließender Silberfärbung von
einigem Material aus diesem zwei-Säulen-gereinigtem Material von
der 1 Liter Zubereitung ergab wenigstens vier bis fünf Banden,
von denen die 10 kD und die 20 kD Banden für dieses Material einzigartig
waren. Passiertes Kulturmedium mit Komponenten mit einem Molekulargewicht
von weniger als 1.000 wurde ohne Verdünnung verwendet.
-
Konzentriertes,
konditioniertes Medium wurde filtersterilisiert mit einem 0,45 μ Uniflo-Filter
(Schleicher und Schuell, Keene, NH) und dann weiter gereinigt durch
Anwendung einer DEAE-SW Anionenaustauschersäule (Waters, Inc., Milford,
MA), die zuvor mit 10 mM Tris-Cl, pH 8,1, preäuquilibriert worden war. Konzentrierte
C. M. Proteine, die 1 Liter des ursprünglichen ATL-2 konditionierten
Medium pro HPLC-Lauf darstellten, wurden an die Säule absorbiert
und dann mit einem linearen Gradienten von 0 mM bis 40 mM NaCl bei
einer Fließrate
von 4 ml/Minute eluiert. Die Fraktionen wurden getestet unter Verwendung
eines in vitro Immunkomplex-Kinasetests mit 10% der geeigneten DEAE-Fraktion
(ein Säulen
gereinigtes Material) oder 1% der geeigneten C18 Fraktion (zwei
Säulen
gereinigtes Material). Die Aktivität, welche die Tyrosinkinaseaktivität von p185c-neu
in einer dosisabhängigen
Weise unter Verwendung des in vitro Immunkomplex-Kinasetests ansteigen ließ, wurde
als ein dominanter Peak über
vier bis fünf
Fraktionen (36–40)
bei etwa 220 bis 240 mM von NaCl eluiert. Nach HPLC-DEAE Reinigung
wurden die Proteine in den aktiven Fraktionen konzentriert und vereinigt,
konzentriert und einer C18 (Millionenmatrix) Umkehrphasenchromatographie
(Waters, Inc. Milford, MA) (bezeichnet als der C18 + 1 Schritt oder
zwei Säulen
gereinigtes Material) unterworfen. Die Elution wurde mit einem linearen
Gradienten von 2-Propanol gegen 0,1% TFA durchgeführt. Alle
Fraktionen wurden gegen RPMI 1640 Medium dialysiert, um das 2-Propanol
zu entfernen, und unter Verwendung des in vitro Immunkomplex-Kinasetests
getestet, der unten beschrieben ist, und eine 1% Konzentration der
geeigneten Fraktion. Die Aktivität,
welche die Tyrosinkinaseaktivität
in p185c-neu ansteigen ließ,
wurde in zwei Peaks eluiert. Ein Peak eluierte in der Fraktion 11–13, während ein
zweiter, etwas weniger aktive Peak in den Fraktionen 20–23 eluierte.
Diese zwei Peaks entsprachen etwa 5 bis 7% Isopropanol beziehungsweise
11 bis 14% Isopropanol. C18#1 erzeugte Fraktionen 11–13 wurden
in den Charakterisierungsstudien verwendet. Die aktiven Fraktionen,
die von dem zweiten Chromatographieschritt erzielt wurden, wurden
zusammengefasst und als die proteinartige Substanzprobe bezeichnet.
-
Eine
20 Liter Zubereitung verwendete die gleiche Reinigungsstrategie.
Die DEAE aktiven Fraktionen 35–41
wurden gesammelt und einer C18 Chromatographie, wie oben diskutiert,
unterworfen. Die C18#1 Fraktionen 11–13 und 21–24 hatten beide eine dosisabhängige Aktivität. Der Pool
der Fraktion 11–13
wurde einem zusätzlichen
C18 Chromatographieschritt unterworfen (bezeichnet als C18#2 oder
drei Säulen
gereinigtes Material). Wiederum hatten die Fraktionen 11–13 und
21–24
die Aktivität.
Die Dosisreaktion der Fraktion 23 wurde durch den in vitro Immunkomplex-Kinasetest, wie im
Beispiel 8 beschrieben, bestimmt und kann erzielt werden durch Zugabe
von 0,005 Volumenprozent an Fraktion 23 und 0,05 Volumenprozent
an Fraktion 23. Dies stellt die größte erzielte Reinheit dar.
-
Die
Molekulargewichtsbereiche wurden auf Basis einer Gelfiltrationschromatographie
und einer Ultrafiltrationsmembrananalyse bestimmt. Nahezu gleiche
Mengen an Tyrosinkinaseaktivität
wurden beibehalten und passierten einen Filter mit einer Absperrung
bei einem Molekulargewicht von 10.000. Fast die gesamte Aktivität passierte
einen Filter mit einer Absperrung bei einem Molekulargewicht von
30.000. Die Molekulargewichtsbereiche für die aktiven chromatographischen
Fraktionen wurden bestimmt durch Vergleich der Fraktionen, welche
die dosierungsabhängige,
neu aktivierende Aktivität
enthielten, mit den Elutionsprofilen eines Satz an Proteinen als
Molekulargewichtsstandards (Sigma Chemical Co., St. Louis, MO),
die unter den gleichen Laufbedingungen erzeugt wurden. Eine Aktivität mit einem
niedrigen Molekulargewicht wurde zwischen 7.000 und 14.000 Daltons
identifiziert. Ein zweiter Bereich an Aktivität ergab sich bei 14.000 bis
etwa 24.000 Daltons.
-
Nach
der Gelelektrophorese unter Verwendung eines Polyacrylamid- Gradientengels (integrated
Separation Systems, Hyde Park, MD oder Phorecase System von Amersham,
Arlington Heights, IL), wurde die Silberfärbung des drei Säulen gereinigten
Materials (C18#2) mit einem kommerziell verfügbaren Silberfärbungskit
(BioRad, Rockville Centre, NY) durchgeführt. Die Fraktionen 21, 22,
23 und 24 von der C18#2 Reinigung der 20 Liter Zubereitung liefen
mit den Markern. Die Fraktionen 22 und 23 zeigten die höchsten Dosierungsreaktionen
in dem 185erbB2 (neu) Kinasetest (siehe
unten). Die Tatsache, dass die ausgewählten Molekulargewichtsfraktionen
mit 185erbB2 interagieren, wurde mit einem
Immunkomplex-Kinasetest demonstriert.
-
Huang
et al. (1992, J. Biol. Chem. 257: 11508–11512), hiermit durch Referenz
eingeführt,
isolierten einen zusätzlichen
neu/erbB2 Ligandenwachstumsfaktor aus der Rinderniere. Der 25 kD
Polypeptidfaktor wurde durch ein Verfahren mit Säulenfraktionierung und anschließender sequenzieller
Säulenchromatographie
auf DEAE/Cellulose (DE52), Sulfadex (sulfatiertes Sephadex G-50),
Heparin-Sepharose 4B und Superdex 75 (schnelle Proteinflüssigchromatographie)
isoliert. Der Faktor, NEL-GF, stimuliert die tyrosinspezifische
Autophosphorilierung des neu/erbB2 Genproduktes.
-
VII. Immunkomplextest
NDF für
die Ligandenbindung an p185erbB2
-
Dieser
Test zeigt die Unterschiede in der Autophosphorilierungsaktivität von immungefälltem p185 durch
die Präinkubation
von PN-NR6 Zelllysat mit verschiedenen Mengen von ATL-2 konditioniertem
Medium (C. H.) oder der proteinartigen Substanz, und wird folgenden
bezeichnet als die neu aktivierende Aktivität.
-
Die
bei dem Immunkomplex-Kinasetest verwendeten Zelllinien wurden erzielt,
hergestellt und kultiviert gemäß den Verfahren,
die offenbart sind in Kokai et al., Cell 55, 287–292 (Juli 28, 1989), deren
Offenbarungen hiermit durch Verweis eingeführt sind, als seien sie selbst
hier aufgeführt,
und in der US-Anmeldung mit der Nummer 386,820, eingereicht am 27.
Juli 1989 im Namen von Mark I. Green, bezeichnet „Methods
of Treating Cancerous Cells with Anti-Receptor Antibodies", deren Offenbarungen
hiermit durch Verweis eingeführt
sind als seien sie vollständig
hier aufgeführt.
-
Die
Zelllinien wurden alle erhalten in DMEM Medium, das 5% FCS enthielt,
als das Kulturmedium (5% FCS-DMEM) bei 37°C in einer angefeuchteten Atmosphäre bei 5%
CO2.
-
Dichte
Kulturen von Zellen in 150 mm Schalen wurden zweimal mit kaltem
PBS gewaschen, in 10 ml gefrorenem-aufgetauten Puffer (150 mM NaCl,
1 mM MgCl2, 20 mM Hepes, pH 7,2, 10% Glycerin,
1 mM EDTA, 1% Aprotinin) gekratzt und zentrifugiert (600 × 6, 10
Minuten). Zellpellets wurden resuspendiert in 1 ml Lysispuffer (50
mM Hepes, pH 7,5, 150 mM NaCl, 3% Brij 35, 1 mM EDTA, 1,5 mM MgCl2, 1% Aprotinin, 1 mM EGTA, 20 μM Na3VO4, 10% Glycerin)
und für
30 Minuten bei 4°C
zentrifugiert. Alle Chemikalien stammten von Sigma Chemical Co.,
St. Louis, Mo, soweit nichts anderes angegeben ist. Das unlösliche Material
wurde durch Zentrifugation bei 40.000 × g für 30 Minuten entfernt. Der
klare Überstand,
der im folgenden verwendet wurde, wird als Zelllysat bezeichnet.
-
Die
Zelllysate wurden für
15 Minuten mit 50 μl
von 50% (Volumen/Volumen) Protein A-Sepharose (Sigma Chemical Co.,
St. Louis, Missouri) inkubiert und für zwei Minuten zentrifugiert,
um die Lysate zu klären.
50 μl Aliquots
der geklärten
Zelllysate wurden auf Eis für
15 Minuten mit konditioniertem Medium, der proteinartigen Substanz
oder anderen Faktoren, wie spezifiziert, in einem Endvolumen von
1 ml mit Lysispuffer inkubiert. Die Proben wurden dann mit 5 μg von 7,164
monoklonalem Antikörper
inkubiert, der die extrazelluläre
Domäne von
p185neu und p185c-neu
erkennt, oder mit anderen geeigneten Antikörpern für 20 Minuten auf Eis inkubiert und
anschließend
erfolgte eine 20 Minuten Inkubation mit 50 μl von 50% (Volumen/Volumen)
Protein A-Sepharose mit Drehung bei 4°C. Immunkomplexe wurden durch
Zentrifugation gesammelt, viermal mit 500 μl Waschpuffer (50 mM Hepes,
pH 7,5, 0,1% Brij 35, 150 mM NaCl, 2 mM EDTA, 1% Aprontinin, 30 μm Na3VO4) gewaschen,
dann zweimal mit Reaktionspuffer (20 mM Hepes (pH 7,4), 3 mM MnCl2 und 0,1% Brij 35, 30 μm Na3VO4). Die Pellets wurden resuspendiert in 50 μl Reaktionspuffer
und (Gamma-33P)-ATP (Amersham, Arlington
Heights, IL) wurde zugegeben, um eine Endkonzentration von 0,2 μm zu erzielen.
Die Proben wurden bei 27°C
für 20
Minuten inkubiert oder bei 4°C
für 25
Minuten mit reineren Proben. Die Reaktionen wurden beendet durch
Zugabe von 3 × SDS
Probenpuffer, der 2 mM ATP und 2 mM EDTA enthielt, und wurden dann
bei 100°C für 5 Minuten
inkubiert. Die Proben wurden dann einer SDS-PAGE Analyse aus 10%
Acrylamidgel unterworfen. Die Gele wurden gefärbt, getrocknet und mit Kodak
XAR oder XRP Film mit Verstärkerfolien
entwickelt.
-
VIII. Reinigung der den
Acetylcholin-Rezeptor induzierenden Aktivität (ARIA)
-
ARIA,
ein 24 kD Protein, welches die Acetylcholin-Rezeptorsynthese stimuliert,
wurde im Labor von Gerald Fischbach (Falls et al., Cell 72: 801–815 (1993))
isoliert. ARIA induziert die Tyrosinphosphorilierung eines 185 kD
Muskelmembranproteins, das p185erbB2 ähnelt und
die Acetylcholinrezeptorsynthese in kultivierten Embryomyotubes
stimuliert. Die Sequenzanalyse der cDNA Klone, die ARIA codieren,
zeigt, dass ARIA ein Mitglied der GGF/erbB2 Ligandengruppe von Proteinen
ist, wobei dies potentiell nützlich
bei der Stimulierung der Gliazellmitogenese und anderen Anwendungen
von zum Beispiel GGF2, wie hier beschrieben, ist.
-
Beispiel 14
-
Proteintyrosinphosphorilierung,
vermittelt durch GGF in Schwann-Zellen
-
Schwann-Zellen
der Ratte zeigen eine Stimulierung der Proteintyrosinphosphorilierung
(36) nach Behandlung mit ausreichenden Mengen an
Gliawachstumsfaktor, um die Proliferation zu induzieren. Unterschiedliche
Mengen an partiell gereinigtem GGF wurden bei einer primären Kultur
von Schwann-Zellen der Ratte gemäß dem Verfahren,
das im Beispiel 3 dargestellt ist, angewandt. Die Schwann-Zellen
wuchsen in DMEM/10% fötales
Kalbsserum/5 μM
Forskolin/0,5 μg
pro ml GGF-CM (0,5 ml pro Vertiefung) in Poly-D-Lysin beschichteten
Platten mit 24 Vertiefungen. Wenn die Zellen zusammenflossen, wurden
die Zellen mit DMEM/10% fötalem
Kälberserum
mit 0,5 ml pro Vertiefung gefüttert
und in einem Inkubator über
Nacht belassen, um sie still zu legen. Am folgenden Tag wurden die
Zellen mit 0,2 ml von DMEM/10% fötalem
Kälberserum gefüttert und
im Inkubator für
1 Stunde belassen. Testproben wurden dann direkt dem Medium mit
verschiedenen Konzentrationen zugesetzt und für verschiedene Zeiträume, wie
erforderlich. Die Zellen wurden dann in kochendem Lysispuffer (Natriumphosphat,
5 mM, pH 6,8; SDS, 2%, β-Mercapteothanol,
5%; Dithiothreitol, 0,1 M; Glycerin, 10%; Bromphenol-Blau, 0,4%;
Natriumvanadat, 10 mM) lysiert, im kochenden Wasserbad für 10 Minuten
inkubiert und dann entweder direkt analysiert oder bei –70°C eingefroren.
-
Die
Proben wurden analysiert durch einen Lauf auf 7,5% SDS-PAGE Gelen
und durch Elektroblotting auf Nitrocellulose unter Verwendung der
Standardverfahren, wie sie beschrieben sind von Towbin et al. (1979) Proc.
Natl. Acad. Sci, USA 76: 4350–4354.
Die geblottete Nitrocellulose wurde mit Antiphosphotyrosin-Antikörper unter
Verwendung von Standardverfahren getestet, wie sie beschrieben sind
in Kamps und Selton (1983), Oncogene 2: 305–315. Die entwickelten Blots
wurden über
Nacht einem Autoradiographiefilm ausgesetzt und unter Verwendung
eines Standardlaborentwicklers entwickelt. Die densiometrischen
Messungen wurden durchgeführt
unter Verwendung eines Ultrascan XL Enhanced Laserdensitometer (LKB).
Die Molekulargewichtsbestimmungen wurden relativ zu vorgefärbten Standards
mit hohem Molekulargewicht (Sigma) gemacht. Die Dosisreaktionen
der Proteinphosphorilierung und der Schwann-Zellproliferationen
waren sehr ähnlich
(36). Das Molekulargewicht der phosphorilierten
Bande ist sehr nahe an dem Molekulargewicht von p185erbB2. Ähnliche
Resultate wurden erzielt, wenn Schwann-Zellen direkt mit konditionierten
Medien behandelt wurden, die von COS-Zellen hergestellt wurden,
die mit dem GGF2HBS5 Klon translatiert worden waren. Diese Ergebnisse
korrelieren sehr gut mit der erwarteten Interaktion der GGFs mit
und der Aktivierung von 185erbB2.
-
Dieses
Experiment wurde mit rekombinantem GGF-II wiederholt. Das konditionierte
Medium, das von einer CHO Zelllinie abstammte, die stabil mit dem
GGF-II Klon (GGF2HBS5) transformiert war, stimulierte die Proteintyrosinphosphorilierung
unter Verwendung des oben beschriebenen Tests. Pseudotransfizierte
CH Zellen stimulierten diese Aktivität nicht ( 52).
-
Beispiel 15
-
Test für die Schwann-Zellproliferation
durch den Proteinfaktor aus der MDA-MB-231 Zelllinie
-
Die
Schwann-Zellproliferation wird vermittelt durch das konditionierte
Medium, das von der menschlichen Brustkrebszelllinie MDA-MB-231
abstammt. Am Tag 1 des Tests wurden 104 primäre Schwann-Zellen
der Ratte in 100 μl
von Dulbecco's modifiziertem
Eagle's Medium gegeben,
das mit 5% fötalem
Rinderplasma pro Vertiefung in einer Mikrotiterplatte mit 96 Vertiefungen
ergänzt
war. Am Tag 2 des Tests wurden 10 μl des konditionierten Mediums
(von der menschlichen Brustkrebszelllinie MDA-MB-231, kultiviert
wie in Beispiel 6 beschrieben) zu jeder Vertiefung der Mikrotiterplatte
zugesetzt. Am Tag 6 wurde die Zahl der Schwann-Zellen pro Platte
bestimmt unter Verwendung eines sauren Phosphatasetests (gemäß dem Verfahren
von Connolly et al. Anal. Biochem. 152: 136 (1986)). Die Schale
wurde mit 100 μl
phosphatgepufferter Kochsalzlösung
(PBS) gewaschen und mit 100 μl
Reaktionspuffer (0,1 M Natriumacetat, (pH 5,5), 0,1% Triton X-100
und 10 mM p-Nitrophenylphosphat) wurde pro Vertiefung zugesetzt.
Die Schale wurde bei 37°C
für 2 Stunden
inkubiert und die Reaktion wurde durch Zugabe von 10 μl von 1 N
NaOH gestoppt. Die optische Dichte von jeder Probe wurde in einem Spektrophotometer
bei 410 mm bestimmt. Eine 38% Stimulation der Zellzahl von Schwann-Zellen, die
mit konditioniertem Medium behandelt waren, gegenüber einer
Kontrollzelllinie (HS-294T, ein Nichtproduzent von den erbB-2 Liganden)
wurde beobachtet. Dieses Ergebnis zeigt, dass ein Protein, das durch
die MDA-MB-231Zelllinie abgegeben wird (welche eine p185erbB2 Bindungsaktivität abgibt), die Schwann-Zellproliferation
stimuliert.
-
Beispiel 16
-
N-Glykosilierung von GGF
-
Die
Proteinsequenz, die von der cDNA Sequenz der GGF-II Kandidatenklone
GGF2BPP1, 2 und 3 abgeleitet ist, enthält eine Reihe an Consensus-N-Glykosilierungsmotiven.
Eine Lücke
in der GGFII02 Peptidsequenz stimmt mit dem Asparaginrest in einem
von diesen Motiven überein,
was anzeigt, dass ein Kohlenhydrat wahrscheinlich an dieser Stelle
gebunden ist.
-
Die
N-Glykosilierung der GGFs wurde untersucht durch Beobachtung der
Mobilitätsveränderungen
auf SDS-PAGE nach Inkubation mit N-Glycanase, was ein Protein ist,
das die kovalenten Bindungen zwischen Kohlenhydraten und Asparaginresten
in Proteinen spaltet.
-
Die
N-Glycanasebehandlung von GGF-II ergab eine Hauptbande bei MW 40–42 kDa
und eine kleinere Bande bei 45–48
kDa. Aktivitätselutionsexperimente
unter nicht-reduzierenden Bedingungen zeigten eine einzelne, aktive
deglykosilierte Spezies von etwa 45–50 kDa.
-
Aktivitätselutionsexperimente
mit GGF-I zeigten auch eine Zunahme der elektrophoretischen Mobilität, wenn
eine Behandlung mit N-Glycanase erfolgte, was eine aktive Spezies
von MW 26–28
kDa ergab. Die Silberfärbung
bestätigte,
dass eine Mobilitätsveränderung
vorlag, obgleich keine N-deglycolisierte
Bande entdeckt werden konnte aufgrund der Hintergrundfärbung in
der verwendeten Probe.
-
Hinterlegung
-
Die
Nukleinsäure,
codierend für
GGF-II (cDNA, GGF2HBS5) Protein (Beispiel 6) in einem Plasmid pBluescript
5k unter der Kontrolle des T7 Promotors wurde hinterlegt bei American
Type Culture Collection, Rockville, Maryland, am 2. September 1992
und erhielt die ATCC Zugangsnummer 75298. Der Anmelder erkennt seine
Verantwortlichkeit hinsichtlich des Ersatzes dieses Plasmides, wenn
dieses vor Ende des Ablaufs des herausgegebenen Patentes nicht lebensfähig werden
sollte, sowie seine Verantwortlichkeit hinsichtlich der Information
des ATCC bei der Erteilung von solch einem Patent, wobei dann die
Hinterlegung der Öffentlichkeit verfügbar gemacht werden
wird. Vor diesem Zeitpunkt wird die Hinterlegung verfügbar sein
durch den „Commissioner
of Patents" gemäß 37 CFR § 1.14 und
35 USC § 112.
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-