CN112074737A - 用于评估李斯特菌菌株的减毒和感染性的组合物和方法 - Google Patents
用于评估李斯特菌菌株的减毒和感染性的组合物和方法 Download PDFInfo
- Publication number
- CN112074737A CN112074737A CN201980029910.3A CN201980029910A CN112074737A CN 112074737 A CN112074737 A CN 112074737A CN 201980029910 A CN201980029910 A CN 201980029910A CN 112074737 A CN112074737 A CN 112074737A
- Authority
- CN
- China
- Prior art keywords
- cells
- listeria
- thp
- lys
- asn
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 241000186781 Listeria Species 0.000 title claims abstract description 182
- 238000000034 method Methods 0.000 title claims abstract description 142
- 239000000203 mixture Substances 0.000 title abstract description 21
- 241000894006 Bacteria Species 0.000 claims abstract description 120
- 241000186779 Listeria monocytogenes Species 0.000 claims abstract description 113
- 210000004027 cell Anatomy 0.000 claims description 310
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 265
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 175
- 101710164436 Listeriolysin O Proteins 0.000 claims description 114
- 239000012634 fragment Substances 0.000 claims description 100
- 230000000890 antigenic effect Effects 0.000 claims description 99
- 208000015181 infectious disease Diseases 0.000 claims description 96
- 150000007523 nucleic acids Chemical class 0.000 claims description 96
- 229920001184 polypeptide Polymers 0.000 claims description 93
- 230000035772 mutation Effects 0.000 claims description 88
- 102000039446 nucleic acids Human genes 0.000 claims description 86
- 108020004707 nucleic acids Proteins 0.000 claims description 86
- 230000004927 fusion Effects 0.000 claims description 83
- 230000001580 bacterial effect Effects 0.000 claims description 73
- 230000012010 growth Effects 0.000 claims description 72
- 239000013612 plasmid Substances 0.000 claims description 71
- 238000012360 testing method Methods 0.000 claims description 70
- 229920001817 Agar Polymers 0.000 claims description 69
- 239000008272 agar Substances 0.000 claims description 69
- 230000002238 attenuated effect Effects 0.000 claims description 54
- 238000012217 deletion Methods 0.000 claims description 53
- 230000037430 deletion Effects 0.000 claims description 53
- PHEDXBVPIONUQT-UHFFFAOYSA-N Cocarcinogen A1 Natural products CCCCCCCCCCCCCC(=O)OC1C(C)C2(O)C3C=C(C)C(=O)C3(O)CC(CO)=CC2C2C1(OC(C)=O)C2(C)C PHEDXBVPIONUQT-UHFFFAOYSA-N 0.000 claims description 43
- 241000607479 Yersinia pestis Species 0.000 claims description 43
- PHEDXBVPIONUQT-RGYGYFBISA-N phorbol 13-acetate 12-myristate Chemical compound C([C@]1(O)C(=O)C(C)=C[C@H]1[C@@]1(O)[C@H](C)[C@H]2OC(=O)CCCCCCCCCCCCC)C(CO)=C[C@H]1[C@H]1[C@]2(OC(C)=O)C1(C)C PHEDXBVPIONUQT-RGYGYFBISA-N 0.000 claims description 42
- 238000010790 dilution Methods 0.000 claims description 39
- 239000012895 dilution Substances 0.000 claims description 39
- 230000003834 intracellular effect Effects 0.000 claims description 38
- 101150023527 actA gene Proteins 0.000 claims description 31
- 210000002540 macrophage Anatomy 0.000 claims description 29
- 230000003115 biocidal effect Effects 0.000 claims description 27
- 108700026244 Open Reading Frames Proteins 0.000 claims description 26
- 101150093386 prfA gene Proteins 0.000 claims description 25
- 101150030499 lnt gene Proteins 0.000 claims description 23
- 229930182566 Gentamicin Natural products 0.000 claims description 22
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 claims description 22
- 229960002518 gentamicin Drugs 0.000 claims description 22
- 101150027417 recU gene Proteins 0.000 claims description 21
- 239000006166 lysate Substances 0.000 claims description 19
- 239000002609 medium Substances 0.000 claims description 19
- 241000701806 Human papillomavirus Species 0.000 claims description 17
- 239000003242 anti bacterial agent Substances 0.000 claims description 15
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 15
- 201000010099 disease Diseases 0.000 claims description 14
- 230000000415 inactivating effect Effects 0.000 claims description 12
- 230000002147 killing effect Effects 0.000 claims description 12
- 230000002934 lysing effect Effects 0.000 claims description 12
- 238000007747 plating Methods 0.000 claims description 12
- 230000001332 colony forming effect Effects 0.000 claims description 11
- 108050004635 D-amino acid aminotransferases Proteins 0.000 claims description 10
- 230000008093 supporting effect Effects 0.000 claims description 10
- 239000001963 growth medium Substances 0.000 claims description 9
- 101001074035 Homo sapiens Zinc finger protein GLI2 Proteins 0.000 claims description 8
- 102100035558 Zinc finger protein GLI2 Human genes 0.000 claims description 8
- 101710132595 Protein E7 Proteins 0.000 claims description 6
- 108010041525 Alanine racemase Proteins 0.000 claims description 5
- 230000000644 propagated effect Effects 0.000 claims description 5
- 239000013074 reference sample Substances 0.000 claims description 5
- 210000004898 n-terminal fragment Anatomy 0.000 claims description 4
- 108090000623 proteins and genes Proteins 0.000 description 239
- 102000004169 proteins and genes Human genes 0.000 description 110
- 235000001014 amino acid Nutrition 0.000 description 109
- 230000015572 biosynthetic process Effects 0.000 description 108
- 229940024606 amino acid Drugs 0.000 description 105
- 235000018102 proteins Nutrition 0.000 description 105
- 150000001413 amino acids Chemical class 0.000 description 104
- 238000003786 synthesis reaction Methods 0.000 description 84
- 238000003556 assay Methods 0.000 description 83
- 101150082952 ACTA1 gene Proteins 0.000 description 66
- 239000000523 sample Substances 0.000 description 49
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 46
- 108020004414 DNA Proteins 0.000 description 42
- 206010028980 Neoplasm Diseases 0.000 description 41
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 40
- 102000004190 Enzymes Human genes 0.000 description 39
- 108090000790 Enzymes Proteins 0.000 description 39
- 229940088598 enzyme Drugs 0.000 description 39
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 36
- 230000002503 metabolic effect Effects 0.000 description 36
- 108010055341 glutamyl-glutamic acid Proteins 0.000 description 34
- 108010076504 Protein Sorting Signals Proteins 0.000 description 32
- 108010029485 Protein Isoforms Proteins 0.000 description 31
- 102000001708 Protein Isoforms Human genes 0.000 description 31
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 29
- 238000006467 substitution reaction Methods 0.000 description 28
- 201000011510 cancer Diseases 0.000 description 25
- WIIZWVCIJKGZOK-RKDXNWHRSA-N chloramphenicol Chemical compound ClC(Cl)C(=O)N[C@H](CO)[C@H](O)C1=CC=C([N+]([O-])=O)C=C1 WIIZWVCIJKGZOK-RKDXNWHRSA-N 0.000 description 24
- 229960005091 chloramphenicol Drugs 0.000 description 23
- 230000014509 gene expression Effects 0.000 description 23
- 229960005322 streptomycin Drugs 0.000 description 23
- 108020004705 Codon Proteins 0.000 description 22
- 125000003275 alpha amino acid group Chemical group 0.000 description 20
- 230000006870 function Effects 0.000 description 20
- 239000002773 nucleotide Substances 0.000 description 20
- 125000003729 nucleotide group Chemical group 0.000 description 20
- 239000013598 vector Substances 0.000 description 20
- 230000035899 viability Effects 0.000 description 20
- 239000000427 antigen Substances 0.000 description 19
- 108091007433 antigens Proteins 0.000 description 19
- 102000036639 antigens Human genes 0.000 description 19
- 239000006285 cell suspension Substances 0.000 description 19
- 101100434723 Bacillus subtilis (strain 168) alr1 gene Proteins 0.000 description 18
- 101100215901 Staphylococcus aureus (strain NCTC 8325 / PS 47) alr2 gene Proteins 0.000 description 18
- 101100171116 Streptococcus pneumoniae (strain ATCC BAA-255 / R6) dprA gene Proteins 0.000 description 18
- 101100276985 Xenopus laevis ccdc88c gene Proteins 0.000 description 18
- 101150070828 alr gene Proteins 0.000 description 18
- 238000011534 incubation Methods 0.000 description 18
- 230000004044 response Effects 0.000 description 18
- 108010006533 ATP-Binding Cassette Transporters Proteins 0.000 description 17
- 102000005416 ATP-Binding Cassette Transporters Human genes 0.000 description 17
- 230000000694 effects Effects 0.000 description 17
- 230000008859 change Effects 0.000 description 16
- 238000009169 immunotherapy Methods 0.000 description 16
- 238000002360 preparation method Methods 0.000 description 16
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 15
- 230000027455 binding Effects 0.000 description 15
- 230000010354 integration Effects 0.000 description 15
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 15
- 101150024289 hly gene Proteins 0.000 description 14
- 238000005259 measurement Methods 0.000 description 14
- SITLTJHOQZFJGG-UHFFFAOYSA-N N-L-alpha-glutamyl-L-valine Natural products CC(C)C(C(O)=O)NC(=O)C(N)CCC(O)=O SITLTJHOQZFJGG-UHFFFAOYSA-N 0.000 description 13
- 108091028043 Nucleic acid sequence Proteins 0.000 description 13
- 210000000680 phagosome Anatomy 0.000 description 13
- 230000001018 virulence Effects 0.000 description 13
- IRXNJYPKBVERCW-DCAQKATOSA-N Glu-Leu-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O IRXNJYPKBVERCW-DCAQKATOSA-N 0.000 description 12
- KDBHVPXBQADZKY-GUBZILKMSA-N Pro-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 KDBHVPXBQADZKY-GUBZILKMSA-N 0.000 description 12
- 108010038633 aspartylglutamate Proteins 0.000 description 12
- 230000028993 immune response Effects 0.000 description 12
- 239000000463 material Substances 0.000 description 12
- 230000001105 regulatory effect Effects 0.000 description 12
- 230000028327 secretion Effects 0.000 description 12
- SBVPYBFMIGDIDX-SRVKXCTJSA-N Pro-Pro-Pro Chemical compound OC(=O)[C@@H]1CCCN1C(=O)[C@H]1N(C(=O)[C@H]2NCCC2)CCC1 SBVPYBFMIGDIDX-SRVKXCTJSA-N 0.000 description 11
- 210000000172 cytosol Anatomy 0.000 description 11
- 108010003700 lysyl aspartic acid Proteins 0.000 description 11
- MJIJBEYEHBKTIM-BYULHYEWSA-N Asn-Val-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N MJIJBEYEHBKTIM-BYULHYEWSA-N 0.000 description 10
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 10
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 10
- 241000880493 Leptailurus serval Species 0.000 description 10
- DNAXXTQSTKOHFO-QEJZJMRPSA-N Phe-Lys-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CC1=CC=CC=C1 DNAXXTQSTKOHFO-QEJZJMRPSA-N 0.000 description 10
- PPCZVWHJWJFTFN-ZLUOBGJFSA-N Ser-Ser-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O PPCZVWHJWJFTFN-ZLUOBGJFSA-N 0.000 description 10
- BMKNXTJLHFIAAH-CIUDSAMLSA-N Ser-Ser-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O BMKNXTJLHFIAAH-CIUDSAMLSA-N 0.000 description 10
- XXWBHOWRARMUOC-NHCYSSNCSA-N Val-Lys-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(=O)N)C(=O)O)N XXWBHOWRARMUOC-NHCYSSNCSA-N 0.000 description 10
- 239000003153 chemical reaction reagent Substances 0.000 description 10
- 238000001802 infusion Methods 0.000 description 10
- 238000003780 insertion Methods 0.000 description 10
- 230000037431 insertion Effects 0.000 description 10
- 239000013641 positive control Substances 0.000 description 10
- 230000008569 process Effects 0.000 description 10
- WDIYWDJLXOCGRW-ACZMJKKPSA-N Ala-Asp-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O WDIYWDJLXOCGRW-ACZMJKKPSA-N 0.000 description 9
- QNAYBMKLOCPYGJ-UWTATZPHSA-N D-alanine Chemical compound C[C@@H](N)C(O)=O QNAYBMKLOCPYGJ-UWTATZPHSA-N 0.000 description 9
- WHUUTDBJXJRKMK-GSVOUGTGSA-N D-glutamic acid Chemical compound OC(=O)[C@H](N)CCC(O)=O WHUUTDBJXJRKMK-GSVOUGTGSA-N 0.000 description 9
- FNGOXVQBBCMFKV-CIUDSAMLSA-N Pro-Ser-Glu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O FNGOXVQBBCMFKV-CIUDSAMLSA-N 0.000 description 9
- 206010038111 Recurrent cancer Diseases 0.000 description 9
- 210000004899 c-terminal region Anatomy 0.000 description 9
- 230000002163 immunogen Effects 0.000 description 9
- 230000005847 immunogenicity Effects 0.000 description 9
- 238000000338 in vitro Methods 0.000 description 9
- 230000010076 replication Effects 0.000 description 9
- 108010061238 threonyl-glycine Proteins 0.000 description 9
- 108010073969 valyllysine Proteins 0.000 description 9
- 102000007469 Actins Human genes 0.000 description 8
- 108010085238 Actins Proteins 0.000 description 8
- 239000004475 Arginine Substances 0.000 description 8
- MGSVBZIBCCKGCY-ZLUOBGJFSA-N Asp-Ser-Ser Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O MGSVBZIBCCKGCY-ZLUOBGJFSA-N 0.000 description 8
- QNAYBMKLOCPYGJ-UHFFFAOYSA-N D-alpha-Ala Natural products CC([NH3+])C([O-])=O QNAYBMKLOCPYGJ-UHFFFAOYSA-N 0.000 description 8
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 8
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 8
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 8
- 108091034117 Oligonucleotide Proteins 0.000 description 8
- MKGIILKDUGDRRO-FXQIFTODSA-N Pro-Ser-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H]1CCCN1 MKGIILKDUGDRRO-FXQIFTODSA-N 0.000 description 8
- 229960003767 alanine Drugs 0.000 description 8
- 235000004279 alanine Nutrition 0.000 description 8
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 8
- 210000004556 brain Anatomy 0.000 description 8
- 238000004422 calculation algorithm Methods 0.000 description 8
- 238000004364 calculation method Methods 0.000 description 8
- 210000000349 chromosome Anatomy 0.000 description 8
- 230000009089 cytolysis Effects 0.000 description 8
- JYPCXBJRLBHWME-UHFFFAOYSA-N glycyl-L-prolyl-L-arginine Natural products NCC(=O)N1CCCC1C(=O)NC(CCCN=C(N)N)C(O)=O JYPCXBJRLBHWME-UHFFFAOYSA-N 0.000 description 8
- 238000001727 in vivo Methods 0.000 description 8
- 108010017391 lysylvaline Proteins 0.000 description 8
- 102000040430 polynucleotide Human genes 0.000 description 8
- 108091033319 polynucleotide Proteins 0.000 description 8
- 239000002157 polynucleotide Substances 0.000 description 8
- 108010077112 prolyl-proline Proteins 0.000 description 8
- 241000894007 species Species 0.000 description 8
- 230000000153 supplemental effect Effects 0.000 description 8
- 238000010257 thawing Methods 0.000 description 8
- 210000001519 tissue Anatomy 0.000 description 8
- HPASIOLTWSNMFB-OLHMAJIHSA-N Asn-Thr-Asp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(O)=O HPASIOLTWSNMFB-OLHMAJIHSA-N 0.000 description 7
- NJIKKGUVGUBICV-ZLUOBGJFSA-N Asp-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC(O)=O NJIKKGUVGUBICV-ZLUOBGJFSA-N 0.000 description 7
- RDDSZZJOKDVPAE-ACZMJKKPSA-N Glu-Asn-Ser Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O RDDSZZJOKDVPAE-ACZMJKKPSA-N 0.000 description 7
- BPCLDCNZBUYGOD-BPUTZDHNSA-N Glu-Trp-Glu Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCC(O)=O)N)C(=O)N[C@@H](CCC(O)=O)C(O)=O)=CNC2=C1 BPCLDCNZBUYGOD-BPUTZDHNSA-N 0.000 description 7
- MTBIKIMYHUWBRX-QWRGUYRKSA-N Gly-Phe-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)CN MTBIKIMYHUWBRX-QWRGUYRKSA-N 0.000 description 7
- JYPCXBJRLBHWME-IUCAKERBSA-N Gly-Pro-Arg Chemical compound NCC(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(O)=O JYPCXBJRLBHWME-IUCAKERBSA-N 0.000 description 7
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 7
- YQFZRHYZLARWDY-IHRRRGAJSA-N Leu-Val-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCCN YQFZRHYZLARWDY-IHRRRGAJSA-N 0.000 description 7
- MPOHDJKRBLVGCT-CIUDSAMLSA-N Lys-Ala-Asn Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCCCN)N MPOHDJKRBLVGCT-CIUDSAMLSA-N 0.000 description 7
- ABHIXYDMILIUKV-CIUDSAMLSA-N Lys-Asn-Asn Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ABHIXYDMILIUKV-CIUDSAMLSA-N 0.000 description 7
- 241001465754 Metazoa Species 0.000 description 7
- OLIJLNWFEQEFDM-SRVKXCTJSA-N Ser-Asp-Phe Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 OLIJLNWFEQEFDM-SRVKXCTJSA-N 0.000 description 7
- LQESNKGTTNHZPZ-GHCJXIJMSA-N Ser-Ile-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O LQESNKGTTNHZPZ-GHCJXIJMSA-N 0.000 description 7
- XHWCDRUPDNSDAZ-XKBZYTNZSA-N Thr-Ser-Glu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N)O XHWCDRUPDNSDAZ-XKBZYTNZSA-N 0.000 description 7
- PVPAOIGJYHVWBT-KKHAAJSZSA-N Val-Asn-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](C(C)C)N)O PVPAOIGJYHVWBT-KKHAAJSZSA-N 0.000 description 7
- 108010087924 alanylproline Proteins 0.000 description 7
- 210000000612 antigen-presenting cell Anatomy 0.000 description 7
- 108010062796 arginyllysine Proteins 0.000 description 7
- 108010092854 aspartyllysine Proteins 0.000 description 7
- 238000011109 contamination Methods 0.000 description 7
- 229940126534 drug product Drugs 0.000 description 7
- 101150046348 inlB gene Proteins 0.000 description 7
- 108010078274 isoleucylvaline Proteins 0.000 description 7
- 239000007788 liquid Substances 0.000 description 7
- 108010054155 lysyllysine Proteins 0.000 description 7
- 239000000825 pharmaceutical preparation Substances 0.000 description 7
- 108010071207 serylmethionine Proteins 0.000 description 7
- 239000008223 sterile water Substances 0.000 description 7
- 238000013518 transcription Methods 0.000 description 7
- 230000035897 transcription Effects 0.000 description 7
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 6
- JVJGCCBAOOWGEO-RUTPOYCXSA-N (2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-4-amino-2-[[(2s,3s)-2-[[(2s,3s)-2-[[(2s)-2-azaniumyl-3-hydroxypropanoyl]amino]-3-methylpentanoyl]amino]-3-methylpentanoyl]amino]-4-oxobutanoyl]amino]-3-phenylpropanoyl]amino]-4-carboxylatobutanoyl]amino]-6-azaniumy Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=CC=C1 JVJGCCBAOOWGEO-RUTPOYCXSA-N 0.000 description 6
- ZIWWTZWAKYBUOB-CIUDSAMLSA-N Ala-Asp-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O ZIWWTZWAKYBUOB-CIUDSAMLSA-N 0.000 description 6
- SAHQGRZIQVEJPF-JXUBOQSCSA-N Ala-Thr-Lys Chemical compound C[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@H](C(O)=O)CCCCN SAHQGRZIQVEJPF-JXUBOQSCSA-N 0.000 description 6
- ACRYGQFHAQHDSF-ZLUOBGJFSA-N Asn-Asn-Asn Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ACRYGQFHAQHDSF-ZLUOBGJFSA-N 0.000 description 6
- PCKRJVZAQZWNKM-WHFBIAKZSA-N Asn-Asn-Gly Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O PCKRJVZAQZWNKM-WHFBIAKZSA-N 0.000 description 6
- KQBVNNAPIURMPD-PEFMBERDSA-N Asp-Ile-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(O)=O KQBVNNAPIURMPD-PEFMBERDSA-N 0.000 description 6
- XQDGOJPVMSWZSO-SRVKXCTJSA-N Gln-Pro-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CCC(=O)N)N XQDGOJPVMSWZSO-SRVKXCTJSA-N 0.000 description 6
- MXOODARRORARSU-ACZMJKKPSA-N Glu-Ala-Ser Chemical compound C[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CCC(=O)O)N MXOODARRORARSU-ACZMJKKPSA-N 0.000 description 6
- HZISRJBYZAODRV-XQXXSGGOSA-N Glu-Thr-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O HZISRJBYZAODRV-XQXXSGGOSA-N 0.000 description 6
- YWAQATDNEKZFFK-BYPYZUCNSA-N Gly-Gly-Ser Chemical compound NCC(=O)NCC(=O)N[C@@H](CO)C(O)=O YWAQATDNEKZFFK-BYPYZUCNSA-N 0.000 description 6
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 6
- RWIKBYVJQAJYDP-BJDJZHNGSA-N Ile-Ala-Lys Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN RWIKBYVJQAJYDP-BJDJZHNGSA-N 0.000 description 6
- WUEIUSDAECDLQO-NAKRPEOUSA-N Ile-Ala-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](CCSC)C(=O)O)N WUEIUSDAECDLQO-NAKRPEOUSA-N 0.000 description 6
- PXKACEXYLPBMAD-JBDRJPRFSA-N Ile-Ser-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)O)N PXKACEXYLPBMAD-JBDRJPRFSA-N 0.000 description 6
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 6
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 6
- JDBQSGMJBMPNFT-AVGNSLFASA-N Leu-Pro-Val Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=O JDBQSGMJBMPNFT-AVGNSLFASA-N 0.000 description 6
- LOGFVTREOLYCPF-RHYQMDGZSA-N Lys-Pro-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCCN LOGFVTREOLYCPF-RHYQMDGZSA-N 0.000 description 6
- NYTDJEZBAAFLLG-IHRRRGAJSA-N Lys-Val-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(O)=O NYTDJEZBAAFLLG-IHRRRGAJSA-N 0.000 description 6
- 108060004795 Methyltransferase Proteins 0.000 description 6
- 108010002311 N-glycylglutamic acid Proteins 0.000 description 6
- OYQBFWWQSVIHBN-FHWLQOOXSA-N Phe-Glu-Phe Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O OYQBFWWQSVIHBN-FHWLQOOXSA-N 0.000 description 6
- 102000007066 Prostate-Specific Antigen Human genes 0.000 description 6
- 108010072866 Prostate-Specific Antigen Proteins 0.000 description 6
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 6
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 6
- KCNSGAMPBPYUAI-CIUDSAMLSA-N Ser-Leu-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O KCNSGAMPBPYUAI-CIUDSAMLSA-N 0.000 description 6
- MUJQWSAWLLRJCE-KATARQTJSA-N Ser-Leu-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O MUJQWSAWLLRJCE-KATARQTJSA-N 0.000 description 6
- DIHPMRTXPYMDJZ-KAOXEZKKSA-N Thr-Tyr-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N2CCC[C@@H]2C(=O)O)N)O DIHPMRTXPYMDJZ-KAOXEZKKSA-N 0.000 description 6
- 241001664469 Tibicina haematodes Species 0.000 description 6
- 238000007792 addition Methods 0.000 description 6
- 238000004458 analytical method Methods 0.000 description 6
- 229940088710 antibiotic agent Drugs 0.000 description 6
- 108010040443 aspartyl-aspartic acid Proteins 0.000 description 6
- 229910002091 carbon monoxide Inorganic materials 0.000 description 6
- 238000004113 cell culture Methods 0.000 description 6
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 6
- 210000000805 cytoplasm Anatomy 0.000 description 6
- 230000037433 frameshift Effects 0.000 description 6
- 235000013922 glutamic acid Nutrition 0.000 description 6
- 239000004220 glutamic acid Substances 0.000 description 6
- 108010082286 glycyl-seryl-alanine Proteins 0.000 description 6
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 6
- 238000011081 inoculation Methods 0.000 description 6
- 229960000310 isoleucine Drugs 0.000 description 6
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 6
- 239000011159 matrix material Substances 0.000 description 6
- 239000013642 negative control Substances 0.000 description 6
- 229960002429 proline Drugs 0.000 description 6
- 108010026333 seryl-proline Proteins 0.000 description 6
- 238000012546 transfer Methods 0.000 description 6
- 230000014616 translation Effects 0.000 description 6
- 229960005486 vaccine Drugs 0.000 description 6
- 238000005406 washing Methods 0.000 description 6
- RRBGTUQJDFBWNN-MUGJNUQGSA-N (2s)-6-amino-2-[[(2s)-6-amino-2-[[(2s)-6-amino-2-[[(2s)-2,6-diaminohexanoyl]amino]hexanoyl]amino]hexanoyl]amino]hexanoic acid Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O RRBGTUQJDFBWNN-MUGJNUQGSA-N 0.000 description 5
- HPYLHFWTUAGUNX-BGZSDMPXSA-N (3s)-4-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-oxo-3-[[(2s,6s)-2,6,10-triamino-4-[(diaminomethylideneamino)methyl]-5-oxodecanoyl]amino]butanoic acid Chemical compound NCCCC[C@H](N)C(=O)C(CN=C(N)N)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O HPYLHFWTUAGUNX-BGZSDMPXSA-N 0.000 description 5
- PIPTUBPKYFRLCP-NHCYSSNCSA-N Ala-Ala-Phe Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 PIPTUBPKYFRLCP-NHCYSSNCSA-N 0.000 description 5
- FXKNPWNXPQZLES-ZLUOBGJFSA-N Ala-Asn-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O FXKNPWNXPQZLES-ZLUOBGJFSA-N 0.000 description 5
- GWFSQQNGMPGBEF-GHCJXIJMSA-N Ala-Asp-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](C)N GWFSQQNGMPGBEF-GHCJXIJMSA-N 0.000 description 5
- HMRWQTHUDVXMGH-GUBZILKMSA-N Ala-Glu-Lys Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CCCCN HMRWQTHUDVXMGH-GUBZILKMSA-N 0.000 description 5
- PNALXAODQKTNLV-JBDRJPRFSA-N Ala-Ile-Ala Chemical compound C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O PNALXAODQKTNLV-JBDRJPRFSA-N 0.000 description 5
- XHNLCGXYBXNRIS-BJDJZHNGSA-N Ala-Lys-Ile Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O XHNLCGXYBXNRIS-BJDJZHNGSA-N 0.000 description 5
- CJQAEJMHBAOQHA-DLOVCJGASA-N Ala-Phe-Asn Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(=O)N)C(=O)O)N CJQAEJMHBAOQHA-DLOVCJGASA-N 0.000 description 5
- IPZQNYYAYVRKKK-FXQIFTODSA-N Ala-Pro-Ala Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O IPZQNYYAYVRKKK-FXQIFTODSA-N 0.000 description 5
- JPOQZCHGOTWRTM-FQPOAREZSA-N Ala-Tyr-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JPOQZCHGOTWRTM-FQPOAREZSA-N 0.000 description 5
- REWSWYIDQIELBE-FXQIFTODSA-N Ala-Val-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O REWSWYIDQIELBE-FXQIFTODSA-N 0.000 description 5
- OMSKGWFGWCQFBD-KZVJFYERSA-N Ala-Val-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OMSKGWFGWCQFBD-KZVJFYERSA-N 0.000 description 5
- NXDXECQFKHXHAM-HJGDQZAQSA-N Arg-Glu-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NXDXECQFKHXHAM-HJGDQZAQSA-N 0.000 description 5
- GOWZVQXTHUCNSQ-NHCYSSNCSA-N Arg-Glu-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O GOWZVQXTHUCNSQ-NHCYSSNCSA-N 0.000 description 5
- LEFKSBYHUGUWLP-ACZMJKKPSA-N Asn-Ala-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O LEFKSBYHUGUWLP-ACZMJKKPSA-N 0.000 description 5
- CMLGVVWQQHUXOZ-GHCJXIJMSA-N Asn-Ala-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O CMLGVVWQQHUXOZ-GHCJXIJMSA-N 0.000 description 5
- QEYJFBMTSMLPKZ-ZKWXMUAHSA-N Asn-Ala-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(O)=O QEYJFBMTSMLPKZ-ZKWXMUAHSA-N 0.000 description 5
- WIDVAWAQBRAKTI-YUMQZZPRSA-N Asn-Leu-Gly Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O WIDVAWAQBRAKTI-YUMQZZPRSA-N 0.000 description 5
- KHCNTVRVAYCPQE-CIUDSAMLSA-N Asn-Lys-Asn Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(O)=O KHCNTVRVAYCPQE-CIUDSAMLSA-N 0.000 description 5
- SNYCNNPOFYBCEK-ZLUOBGJFSA-N Asn-Ser-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O SNYCNNPOFYBCEK-ZLUOBGJFSA-N 0.000 description 5
- JBDLMLZNDRLDIX-HJGDQZAQSA-N Asn-Thr-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O JBDLMLZNDRLDIX-HJGDQZAQSA-N 0.000 description 5
- CBWCQCANJSGUOH-ZKWXMUAHSA-N Asn-Val-Ala Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O CBWCQCANJSGUOH-ZKWXMUAHSA-N 0.000 description 5
- CBHVAFXKOYAHOY-NHCYSSNCSA-N Asn-Val-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O CBHVAFXKOYAHOY-NHCYSSNCSA-N 0.000 description 5
- PQKSVQSMTHPRIB-ZKWXMUAHSA-N Asn-Val-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O PQKSVQSMTHPRIB-ZKWXMUAHSA-N 0.000 description 5
- RGKKALNPOYURGE-ZKWXMUAHSA-N Asp-Ala-Val Chemical compound N[C@@H](CC(=O)O)C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)O RGKKALNPOYURGE-ZKWXMUAHSA-N 0.000 description 5
- SCQIQCWLOMOEFP-DCAQKATOSA-N Asp-Leu-Arg Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O SCQIQCWLOMOEFP-DCAQKATOSA-N 0.000 description 5
- USENATHVGFXRNO-SRVKXCTJSA-N Asp-Tyr-Asp Chemical compound OC(=O)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(O)=O)C(O)=O)CC1=CC=C(O)C=C1 USENATHVGFXRNO-SRVKXCTJSA-N 0.000 description 5
- XMKXONRMGJXCJV-LAEOZQHASA-N Asp-Val-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O XMKXONRMGJXCJV-LAEOZQHASA-N 0.000 description 5
- 108700010070 Codon Usage Proteins 0.000 description 5
- SOBBAYVQSNXYPQ-ACZMJKKPSA-N Gln-Asn-Asn Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O SOBBAYVQSNXYPQ-ACZMJKKPSA-N 0.000 description 5
- QYTKAVBFRUGYAU-ACZMJKKPSA-N Gln-Asp-Asn Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O QYTKAVBFRUGYAU-ACZMJKKPSA-N 0.000 description 5
- NPTGGVQJYRSMCM-GLLZPBPUSA-N Gln-Gln-Thr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NPTGGVQJYRSMCM-GLLZPBPUSA-N 0.000 description 5
- JKGHMESJHRTHIC-SIUGBPQLSA-N Gln-Ile-Tyr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)NC(=O)[C@H](CCC(=O)N)N JKGHMESJHRTHIC-SIUGBPQLSA-N 0.000 description 5
- CSMHMEATMDCQNY-DZKIICNBSA-N Gln-Val-Tyr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O CSMHMEATMDCQNY-DZKIICNBSA-N 0.000 description 5
- SOEXCCGNHQBFPV-DLOVCJGASA-N Gln-Val-Val Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C(C)C)C(O)=O SOEXCCGNHQBFPV-DLOVCJGASA-N 0.000 description 5
- JJKKWYQVHRUSDG-GUBZILKMSA-N Glu-Ala-Lys Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O JJKKWYQVHRUSDG-GUBZILKMSA-N 0.000 description 5
- MLCPTRRNICEKIS-FXQIFTODSA-N Glu-Asn-Gln Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O MLCPTRRNICEKIS-FXQIFTODSA-N 0.000 description 5
- HNVFSTLPVJWIDV-CIUDSAMLSA-N Glu-Glu-Gln Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O HNVFSTLPVJWIDV-CIUDSAMLSA-N 0.000 description 5
- LGYZYFFDELZWRS-DCAQKATOSA-N Glu-Glu-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O LGYZYFFDELZWRS-DCAQKATOSA-N 0.000 description 5
- KRGZZKWSBGPLKL-IUCAKERBSA-N Glu-Gly-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CCC(=O)O)N KRGZZKWSBGPLKL-IUCAKERBSA-N 0.000 description 5
- GJBUAAAIZSRCDC-GVXVVHGQSA-N Glu-Leu-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O GJBUAAAIZSRCDC-GVXVVHGQSA-N 0.000 description 5
- ZQYZDDXTNQXUJH-CIUDSAMLSA-N Glu-Met-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CCC(=O)O)N ZQYZDDXTNQXUJH-CIUDSAMLSA-N 0.000 description 5
- HJTSRYLPAYGEEC-SIUGBPQLSA-N Glu-Tyr-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)NC(=O)[C@H](CCC(=O)O)N HJTSRYLPAYGEEC-SIUGBPQLSA-N 0.000 description 5
- FGGKGJHCVMYGCD-UKJIMTQDSA-N Glu-Val-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O FGGKGJHCVMYGCD-UKJIMTQDSA-N 0.000 description 5
- YMUFWNJHVPQNQD-ZKWXMUAHSA-N Gly-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)CN YMUFWNJHVPQNQD-ZKWXMUAHSA-N 0.000 description 5
- NTBOEZICHOSJEE-YUMQZZPRSA-N Gly-Lys-Ser Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(O)=O NTBOEZICHOSJEE-YUMQZZPRSA-N 0.000 description 5
- SCJJPCQUJYPHRZ-BQBZGAKWSA-N Gly-Pro-Asn Chemical compound NCC(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O SCJJPCQUJYPHRZ-BQBZGAKWSA-N 0.000 description 5
- DNVDEMWIYLVIQU-RCOVLWMOSA-N Gly-Val-Asp Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O DNVDEMWIYLVIQU-RCOVLWMOSA-N 0.000 description 5
- ILUVWFTXAUYOBW-CUJWVEQBSA-N His-Ser-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC1=CN=CN1)N)O ILUVWFTXAUYOBW-CUJWVEQBSA-N 0.000 description 5
- SCHZQZPYHBWYEQ-PEFMBERDSA-N Ile-Asn-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N SCHZQZPYHBWYEQ-PEFMBERDSA-N 0.000 description 5
- NKRJALPCDNXULF-BYULHYEWSA-N Ile-Asp-Gly Chemical compound [H]N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O NKRJALPCDNXULF-BYULHYEWSA-N 0.000 description 5
- QSPLUJGYOPZINY-ZPFDUUQYSA-N Ile-Asp-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N QSPLUJGYOPZINY-ZPFDUUQYSA-N 0.000 description 5
- CYHJCEKUMCNDFG-LAEOZQHASA-N Ile-Gln-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)NCC(=O)O)N CYHJCEKUMCNDFG-LAEOZQHASA-N 0.000 description 5
- PNDMHTTXXPUQJH-RWRJDSDZSA-N Ile-Glu-Thr Chemical compound N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H]([C@H](O)C)C(=O)O PNDMHTTXXPUQJH-RWRJDSDZSA-N 0.000 description 5
- CSQNHSGHAPRGPQ-YTFOTSKYSA-N Ile-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCCN)C(=O)O)N CSQNHSGHAPRGPQ-YTFOTSKYSA-N 0.000 description 5
- TVYWVSJGSHQWMT-AJNGGQMLSA-N Ile-Leu-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)O)N TVYWVSJGSHQWMT-AJNGGQMLSA-N 0.000 description 5
- WSSGUVAKYCQSCT-XUXIUFHCSA-N Ile-Met-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(=O)O)N WSSGUVAKYCQSCT-XUXIUFHCSA-N 0.000 description 5
- 108010061833 Integrases Proteins 0.000 description 5
- PMGDADKJMCOXHX-UHFFFAOYSA-N L-Arginyl-L-glutamin-acetat Natural products NC(=N)NCCCC(N)C(=O)NC(CCC(N)=O)C(O)=O PMGDADKJMCOXHX-UHFFFAOYSA-N 0.000 description 5
- QLQHWWCSCLZUMA-KKUMJFAQSA-N Leu-Asp-Tyr Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 QLQHWWCSCLZUMA-KKUMJFAQSA-N 0.000 description 5
- VQPPIMUZCZCOIL-GUBZILKMSA-N Leu-Gln-Ala Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O VQPPIMUZCZCOIL-GUBZILKMSA-N 0.000 description 5
- POZULHZYLPGXMR-ONGXEEELSA-N Leu-Gly-Val Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O POZULHZYLPGXMR-ONGXEEELSA-N 0.000 description 5
- UCBPDSYUVAAHCD-UWVGGRQHSA-N Leu-Pro-Gly Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O UCBPDSYUVAAHCD-UWVGGRQHSA-N 0.000 description 5
- AEDWWMMHUGYIFD-HJGDQZAQSA-N Leu-Thr-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(O)=O AEDWWMMHUGYIFD-HJGDQZAQSA-N 0.000 description 5
- QWWPYKKLXWOITQ-VOAKCMCISA-N Leu-Thr-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC(C)C QWWPYKKLXWOITQ-VOAKCMCISA-N 0.000 description 5
- VKVDRTGWLVZJOM-DCAQKATOSA-N Leu-Val-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O VKVDRTGWLVZJOM-DCAQKATOSA-N 0.000 description 5
- OVIVOCSURJYCTM-GUBZILKMSA-N Lys-Asp-Glu Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCC(O)=O OVIVOCSURJYCTM-GUBZILKMSA-N 0.000 description 5
- AAORVPFVUIHEAB-YUMQZZPRSA-N Lys-Asp-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O AAORVPFVUIHEAB-YUMQZZPRSA-N 0.000 description 5
- GRADYHMSAUIKPS-DCAQKATOSA-N Lys-Glu-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O GRADYHMSAUIKPS-DCAQKATOSA-N 0.000 description 5
- ITWQLSZTLBKWJM-YUMQZZPRSA-N Lys-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)[C@@H](N)CCCCN ITWQLSZTLBKWJM-YUMQZZPRSA-N 0.000 description 5
- NNKLKUUGESXCBS-KBPBESRZSA-N Lys-Gly-Tyr Chemical compound [H]N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O NNKLKUUGESXCBS-KBPBESRZSA-N 0.000 description 5
- QOJDBRUCOXQSSK-AJNGGQMLSA-N Lys-Ile-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCCN)C(O)=O QOJDBRUCOXQSSK-AJNGGQMLSA-N 0.000 description 5
- WAIHHELKYSFIQN-XUXIUFHCSA-N Lys-Ile-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O WAIHHELKYSFIQN-XUXIUFHCSA-N 0.000 description 5
- WRODMZBHNNPRLN-SRVKXCTJSA-N Lys-Leu-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O WRODMZBHNNPRLN-SRVKXCTJSA-N 0.000 description 5
- JQSIGLHQNSZZRL-KKUMJFAQSA-N Lys-Lys-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCCCN)N JQSIGLHQNSZZRL-KKUMJFAQSA-N 0.000 description 5
- 239000004472 Lysine Substances 0.000 description 5
- FWTBMGAKKPSTBT-GUBZILKMSA-N Met-Gln-Glu Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O FWTBMGAKKPSTBT-GUBZILKMSA-N 0.000 description 5
- HAQLBBVZAGMESV-IHRRRGAJSA-N Met-Lys-Lys Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O HAQLBBVZAGMESV-IHRRRGAJSA-N 0.000 description 5
- RIIFMEBFDDXGCV-VEVYYDQMSA-N Met-Thr-Asn Chemical compound CSCC[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC(N)=O RIIFMEBFDDXGCV-VEVYYDQMSA-N 0.000 description 5
- YBAFDPFAUTYYRW-UHFFFAOYSA-N N-L-alpha-glutamyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=O YBAFDPFAUTYYRW-UHFFFAOYSA-N 0.000 description 5
- QPVFUAUFEBPIPT-CDMKHQONSA-N Phe-Gly-Thr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(O)=O QPVFUAUFEBPIPT-CDMKHQONSA-N 0.000 description 5
- IWZRODDWOSIXPZ-IRXDYDNUSA-N Phe-Phe-Gly Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)NCC(O)=O)C1=CC=CC=C1 IWZRODDWOSIXPZ-IRXDYDNUSA-N 0.000 description 5
- XQLBWXHVZVBNJM-FXQIFTODSA-N Pro-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 XQLBWXHVZVBNJM-FXQIFTODSA-N 0.000 description 5
- XUSDDSLCRPUKLP-QXEWZRGKSA-N Pro-Asp-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H]1CCCN1 XUSDDSLCRPUKLP-QXEWZRGKSA-N 0.000 description 5
- FKLSMYYLJHYPHH-UWVGGRQHSA-N Pro-Gly-Leu Chemical compound [H]N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O FKLSMYYLJHYPHH-UWVGGRQHSA-N 0.000 description 5
- HAEGAELAYWSUNC-WPRPVWTQSA-N Pro-Gly-Val Chemical compound [H]N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O HAEGAELAYWSUNC-WPRPVWTQSA-N 0.000 description 5
- IBGCFJDLCYTKPW-NAKRPEOUSA-N Pro-Ile-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H]1CCCN1 IBGCFJDLCYTKPW-NAKRPEOUSA-N 0.000 description 5
- KWMUAKQOVYCQJQ-ZPFDUUQYSA-N Pro-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@@H]1CCCN1 KWMUAKQOVYCQJQ-ZPFDUUQYSA-N 0.000 description 5
- WFIVLLFYUZZWOD-RHYQMDGZSA-N Pro-Lys-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O WFIVLLFYUZZWOD-RHYQMDGZSA-N 0.000 description 5
- JLMZKEQFMVORMA-SRVKXCTJSA-N Pro-Pro-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 JLMZKEQFMVORMA-SRVKXCTJSA-N 0.000 description 5
- GOMUXSCOIWIJFP-GUBZILKMSA-N Pro-Ser-Arg Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O GOMUXSCOIWIJFP-GUBZILKMSA-N 0.000 description 5
- PKHDJFHFMGQMPS-RCWTZXSCSA-N Pro-Thr-Arg Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O PKHDJFHFMGQMPS-RCWTZXSCSA-N 0.000 description 5
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 5
- BTKUIVBNGBFTTP-WHFBIAKZSA-N Ser-Ala-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)NCC(O)=O BTKUIVBNGBFTTP-WHFBIAKZSA-N 0.000 description 5
- OJPHFSOMBZKQKQ-GUBZILKMSA-N Ser-Gln-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](N)CO OJPHFSOMBZKQKQ-GUBZILKMSA-N 0.000 description 5
- HJEBZBMOTCQYDN-ACZMJKKPSA-N Ser-Glu-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O HJEBZBMOTCQYDN-ACZMJKKPSA-N 0.000 description 5
- UOLGINIHBRIECN-FXQIFTODSA-N Ser-Glu-Glu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O UOLGINIHBRIECN-FXQIFTODSA-N 0.000 description 5
- WBINSDOPZHQPPM-AVGNSLFASA-N Ser-Glu-Tyr Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CO)N)O WBINSDOPZHQPPM-AVGNSLFASA-N 0.000 description 5
- BKZYBLLIBOBOOW-GHCJXIJMSA-N Ser-Ile-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O BKZYBLLIBOBOOW-GHCJXIJMSA-N 0.000 description 5
- XVWDJUROVRQKAE-KKUMJFAQSA-N Ser-Phe-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)CO)CC1=CC=CC=C1 XVWDJUROVRQKAE-KKUMJFAQSA-N 0.000 description 5
- KQNDIKOYWZTZIX-FXQIFTODSA-N Ser-Ser-Arg Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCNC(N)=N KQNDIKOYWZTZIX-FXQIFTODSA-N 0.000 description 5
- ZSDXEKUKQAKZFE-XAVMHZPKSA-N Ser-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N)O ZSDXEKUKQAKZFE-XAVMHZPKSA-N 0.000 description 5
- PMTWIUBUQRGCSB-FXQIFTODSA-N Ser-Val-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O PMTWIUBUQRGCSB-FXQIFTODSA-N 0.000 description 5
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 5
- 230000005867 T cell response Effects 0.000 description 5
- 210000001744 T-lymphocyte Anatomy 0.000 description 5
- JTEICXDKGWKRRV-HJGDQZAQSA-N Thr-Asn-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N)O JTEICXDKGWKRRV-HJGDQZAQSA-N 0.000 description 5
- OJRNZRROAIAHDL-LKXGYXEUSA-N Thr-Asn-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O OJRNZRROAIAHDL-LKXGYXEUSA-N 0.000 description 5
- PZVGOVRNGKEFCB-KKHAAJSZSA-N Thr-Asn-Val Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](C(C)C)C(=O)O)N)O PZVGOVRNGKEFCB-KKHAAJSZSA-N 0.000 description 5
- FLPZMPOZGYPBEN-PPCPHDFISA-N Thr-Leu-Ile Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O FLPZMPOZGYPBEN-PPCPHDFISA-N 0.000 description 5
- KZURUCDWKDEAFZ-XVSYOHENSA-N Thr-Phe-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)O KZURUCDWKDEAFZ-XVSYOHENSA-N 0.000 description 5
- IQPWNQRRAJHOKV-KATARQTJSA-N Thr-Ser-Lys Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCCN IQPWNQRRAJHOKV-KATARQTJSA-N 0.000 description 5
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 5
- 239000004473 Threonine Substances 0.000 description 5
- IXEGQBJZDIRRIV-QEJZJMRPSA-N Trp-Asn-Glu Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O IXEGQBJZDIRRIV-QEJZJMRPSA-N 0.000 description 5
- XLMDWQNAOKLKCP-XDTLVQLUSA-N Tyr-Ala-Gln Chemical compound C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N XLMDWQNAOKLKCP-XDTLVQLUSA-N 0.000 description 5
- PMDWYLVWHRTJIW-STQMWFEESA-N Tyr-Gly-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC1=CC=C(O)C=C1 PMDWYLVWHRTJIW-STQMWFEESA-N 0.000 description 5
- QARCDOCCDOLJSF-HJPIBITLSA-N Tyr-Ile-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N QARCDOCCDOLJSF-HJPIBITLSA-N 0.000 description 5
- QFXVAFIHVWXXBJ-AVGNSLFASA-N Tyr-Ser-Glu Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O QFXVAFIHVWXXBJ-AVGNSLFASA-N 0.000 description 5
- BYOHPUZJVXWHAE-BYULHYEWSA-N Val-Asn-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC(=O)N)C(=O)O)N BYOHPUZJVXWHAE-BYULHYEWSA-N 0.000 description 5
- ZMDCGGKHRKNWKD-LAEOZQHASA-N Val-Asn-Glu Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N ZMDCGGKHRKNWKD-LAEOZQHASA-N 0.000 description 5
- QGFPYRPIUXBYGR-YDHLFZDLSA-N Val-Asn-Phe Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N QGFPYRPIUXBYGR-YDHLFZDLSA-N 0.000 description 5
- CPTQYHDSVGVGDZ-UKJIMTQDSA-N Val-Gln-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](C(C)C)N CPTQYHDSVGVGDZ-UKJIMTQDSA-N 0.000 description 5
- BRPKEERLGYNCNC-NHCYSSNCSA-N Val-Glu-Arg Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N BRPKEERLGYNCNC-NHCYSSNCSA-N 0.000 description 5
- BZWUSZGQOILYEU-STECZYCISA-N Val-Ile-Tyr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 BZWUSZGQOILYEU-STECZYCISA-N 0.000 description 5
- FMQGYTMERWBMSI-HJWJTTGWSA-N Val-Phe-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)[C@H](C(C)C)N FMQGYTMERWBMSI-HJWJTTGWSA-N 0.000 description 5
- MHHAWNPHDLCPLF-ULQDDVLXSA-N Val-Phe-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)CC1=CC=CC=C1 MHHAWNPHDLCPLF-ULQDDVLXSA-N 0.000 description 5
- UGFMVXRXULGLNO-XPUUQOCRSA-N Val-Ser-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O UGFMVXRXULGLNO-XPUUQOCRSA-N 0.000 description 5
- JPBGMZDTPVGGMQ-ULQDDVLXSA-N Val-Tyr-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N JPBGMZDTPVGGMQ-ULQDDVLXSA-N 0.000 description 5
- NLNCNKIVJPEFBC-DLOVCJGASA-N Val-Val-Glu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCC(O)=O NLNCNKIVJPEFBC-DLOVCJGASA-N 0.000 description 5
- 108010086434 alanyl-seryl-glycine Proteins 0.000 description 5
- 108010005233 alanylglutamic acid Proteins 0.000 description 5
- 108010044940 alanylglutamine Proteins 0.000 description 5
- 125000000539 amino acid group Chemical group 0.000 description 5
- 108010008355 arginyl-glutamine Proteins 0.000 description 5
- 108010060035 arginylproline Proteins 0.000 description 5
- 230000008827 biological function Effects 0.000 description 5
- 238000000423 cell based assay Methods 0.000 description 5
- 230000001413 cellular effect Effects 0.000 description 5
- 238000006243 chemical reaction Methods 0.000 description 5
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 5
- 235000018417 cysteine Nutrition 0.000 description 5
- 230000006378 damage Effects 0.000 description 5
- 238000001514 detection method Methods 0.000 description 5
- 231100000221 frame shift mutation induction Toxicity 0.000 description 5
- 210000004602 germ cell Anatomy 0.000 description 5
- 108010090037 glycyl-alanyl-isoleucine Proteins 0.000 description 5
- 108010059898 glycyl-tyrosyl-lysine Proteins 0.000 description 5
- 230000002949 hemolytic effect Effects 0.000 description 5
- 108010040030 histidinoalanine Proteins 0.000 description 5
- 108010036413 histidylglycine Proteins 0.000 description 5
- 230000001965 increasing effect Effects 0.000 description 5
- 235000018977 lysine Nutrition 0.000 description 5
- 108010025153 lysyl-alanyl-alanine Proteins 0.000 description 5
- 108010064235 lysylglycine Proteins 0.000 description 5
- 108010038320 lysylphenylalanine Proteins 0.000 description 5
- 238000004519 manufacturing process Methods 0.000 description 5
- 238000002156 mixing Methods 0.000 description 5
- 235000015097 nutrients Nutrition 0.000 description 5
- 108010084525 phenylalanyl-phenylalanyl-glycine Proteins 0.000 description 5
- 101150050662 plcB gene Proteins 0.000 description 5
- 108010093296 prolyl-prolyl-alanine Proteins 0.000 description 5
- 230000002829 reductive effect Effects 0.000 description 5
- 230000000717 retained effect Effects 0.000 description 5
- 238000013207 serial dilution Methods 0.000 description 5
- 108010048818 seryl-histidine Proteins 0.000 description 5
- 239000000126 substance Substances 0.000 description 5
- 239000006228 supernatant Substances 0.000 description 5
- 108010031491 threonyl-lysyl-glutamic acid Proteins 0.000 description 5
- 108010072986 threonyl-seryl-lysine Proteins 0.000 description 5
- 108010015385 valyl-prolyl-proline Proteins 0.000 description 5
- GMKMEZVLHJARHF-UHFFFAOYSA-N (2R,6R)-form-2.6-Diaminoheptanedioic acid Natural products OC(=O)C(N)CCCC(N)C(O)=O GMKMEZVLHJARHF-UHFFFAOYSA-N 0.000 description 4
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 4
- JPGBXANAQYHTLA-DRZSPHRISA-N Ala-Gln-Phe Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 JPGBXANAQYHTLA-DRZSPHRISA-N 0.000 description 4
- FFZJHQODAYHGPO-KZVJFYERSA-N Ala-Pro-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](C)N FFZJHQODAYHGPO-KZVJFYERSA-N 0.000 description 4
- YNOCMHZSWJMGBB-GCJQMDKQSA-N Ala-Thr-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(O)=O YNOCMHZSWJMGBB-GCJQMDKQSA-N 0.000 description 4
- JGDGLDNAQJJGJI-AVGNSLFASA-N Arg-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCCN=C(N)N)N JGDGLDNAQJJGJI-AVGNSLFASA-N 0.000 description 4
- HJVGMOYJDDXLMI-AVGNSLFASA-N Arg-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H](N)CCCNC(N)=N HJVGMOYJDDXLMI-AVGNSLFASA-N 0.000 description 4
- ANPFQTJEPONRPL-UGYAYLCHSA-N Asn-Ile-Asp Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O ANPFQTJEPONRPL-UGYAYLCHSA-N 0.000 description 4
- SKQTXVZTCGSRJS-SRVKXCTJSA-N Asn-Tyr-Asp Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CC(=O)N)N)O SKQTXVZTCGSRJS-SRVKXCTJSA-N 0.000 description 4
- XJQRWGXKUSDEFI-ACZMJKKPSA-N Asp-Glu-Asn Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O XJQRWGXKUSDEFI-ACZMJKKPSA-N 0.000 description 4
- YDJVIBMKAMQPPP-LAEOZQHASA-N Asp-Glu-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O YDJVIBMKAMQPPP-LAEOZQHASA-N 0.000 description 4
- 101100343501 Bacillus subtilis (strain 168) lipO gene Proteins 0.000 description 4
- 108091026890 Coding region Proteins 0.000 description 4
- 229930195713 D-glutamate Natural products 0.000 description 4
- 229930182847 D-glutamic acid Natural products 0.000 description 4
- ULGZDMOVFRHVEP-RWJQBGPGSA-N Erythromycin Chemical compound O([C@@H]1[C@@H](C)C(=O)O[C@@H]([C@@]([C@H](O)[C@@H](C)C(=O)[C@H](C)C[C@@](C)(O)[C@H](O[C@H]2[C@@H]([C@H](C[C@@H](C)O2)N(C)C)O)[C@H]1C)(C)O)CC)[C@H]1C[C@@](C)(OC)[C@@H](O)[C@H](C)O1 ULGZDMOVFRHVEP-RWJQBGPGSA-N 0.000 description 4
- XFAUJGNLHIGXET-AVGNSLFASA-N Gln-Leu-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O XFAUJGNLHIGXET-AVGNSLFASA-N 0.000 description 4
- ININBLZFFVOQIO-JHEQGTHGSA-N Gln-Thr-Gly Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CCC(=O)N)N)O ININBLZFFVOQIO-JHEQGTHGSA-N 0.000 description 4
- BBFCMGBMYIAGRS-AUTRQRHGSA-N Gln-Val-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O BBFCMGBMYIAGRS-AUTRQRHGSA-N 0.000 description 4
- QDMVXRNLOPTPIE-WDCWCFNPSA-N Glu-Lys-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QDMVXRNLOPTPIE-WDCWCFNPSA-N 0.000 description 4
- UMZHHILWZBFPGL-LOKLDPHHSA-N Glu-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)O)N)O UMZHHILWZBFPGL-LOKLDPHHSA-N 0.000 description 4
- KKBWDNZXYLGJEY-UHFFFAOYSA-N Gly-Arg-Pro Natural products NCC(=O)NC(CCNC(=N)N)C(=O)N1CCCC1C(=O)O KKBWDNZXYLGJEY-UHFFFAOYSA-N 0.000 description 4
- PTIIBFKSLCYQBO-NHCYSSNCSA-N Gly-Lys-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)CN PTIIBFKSLCYQBO-NHCYSSNCSA-N 0.000 description 4
- DUAWRXXTOQOECJ-JSGCOSHPSA-N Gly-Tyr-Val Chemical compound [H]NCC(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(O)=O DUAWRXXTOQOECJ-JSGCOSHPSA-N 0.000 description 4
- ZHHLTWUOWXHVQJ-YUMQZZPRSA-N His-Ser-Gly Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CO)C(=O)NCC(=O)O)N ZHHLTWUOWXHVQJ-YUMQZZPRSA-N 0.000 description 4
- GVNNAHIRSDRIII-AJNGGQMLSA-N Ile-Lys-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N GVNNAHIRSDRIII-AJNGGQMLSA-N 0.000 description 4
- JDCQDJVYUXNCGF-SPOWBLRKSA-N Ile-Ser-Trp Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N JDCQDJVYUXNCGF-SPOWBLRKSA-N 0.000 description 4
- 102100034343 Integrase Human genes 0.000 description 4
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 4
- 229930182816 L-glutamine Natural products 0.000 description 4
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 4
- KOSWSHVQIVTVQF-ZPFDUUQYSA-N Leu-Ile-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O KOSWSHVQIVTVQF-ZPFDUUQYSA-N 0.000 description 4
- 108090000364 Ligases Proteins 0.000 description 4
- 102000003960 Ligases Human genes 0.000 description 4
- 241000866438 Listeria monocytogenes 10403S Species 0.000 description 4
- WSXTWLJHTLRFLW-SRVKXCTJSA-N Lys-Ala-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O WSXTWLJHTLRFLW-SRVKXCTJSA-N 0.000 description 4
- XOQMURBBIXRRCR-SRVKXCTJSA-N Lys-Lys-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCCN XOQMURBBIXRRCR-SRVKXCTJSA-N 0.000 description 4
- LECIJRIRMVOFMH-ULQDDVLXSA-N Lys-Pro-Phe Chemical compound NCCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 LECIJRIRMVOFMH-ULQDDVLXSA-N 0.000 description 4
- RZJOHSFAEZBWLK-CIUDSAMLSA-N Met-Gln-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CO)C(=O)O)N RZJOHSFAEZBWLK-CIUDSAMLSA-N 0.000 description 4
- ZVRJWDUPIDMHDN-ULQDDVLXSA-N Phe-Pro-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CC1=CC=CC=C1 ZVRJWDUPIDMHDN-ULQDDVLXSA-N 0.000 description 4
- VPEVBAUSTBWQHN-NHCYSSNCSA-N Pro-Glu-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O VPEVBAUSTBWQHN-NHCYSSNCSA-N 0.000 description 4
- VGNYHOBZJKWRGI-CIUDSAMLSA-N Ser-Asn-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CO VGNYHOBZJKWRGI-CIUDSAMLSA-N 0.000 description 4
- UDQBCBUXAQIZAK-GLLZPBPUSA-N Thr-Glu-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O UDQBCBUXAQIZAK-GLLZPBPUSA-N 0.000 description 4
- MBLJBGZWLHTJBH-SZMVWBNQSA-N Trp-Val-Arg Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O)=CNC2=C1 MBLJBGZWLHTJBH-SZMVWBNQSA-N 0.000 description 4
- XJPXTYLVMUZGNW-IHRRRGAJSA-N Tyr-Pro-Asp Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O XJPXTYLVMUZGNW-IHRRRGAJSA-N 0.000 description 4
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 4
- 230000037354 amino acid metabolism Effects 0.000 description 4
- 230000030741 antigen processing and presentation Effects 0.000 description 4
- 101150037081 aroA gene Proteins 0.000 description 4
- 108010077245 asparaginyl-proline Proteins 0.000 description 4
- 230000008901 benefit Effects 0.000 description 4
- 230000004071 biological effect Effects 0.000 description 4
- 230000007423 decrease Effects 0.000 description 4
- 230000004069 differentiation Effects 0.000 description 4
- 238000009472 formulation Methods 0.000 description 4
- 108010062266 glycyl-glycyl-argininal Proteins 0.000 description 4
- 108010010096 glycyl-glycyl-tyrosine Proteins 0.000 description 4
- 108010085325 histidylproline Proteins 0.000 description 4
- 230000001506 immunosuppresive effect Effects 0.000 description 4
- 108010051673 leucyl-glycyl-phenylalanine Proteins 0.000 description 4
- 101150052914 lolA gene Proteins 0.000 description 4
- 101150095537 lplA gene Proteins 0.000 description 4
- 238000012423 maintenance Methods 0.000 description 4
- 239000003550 marker Substances 0.000 description 4
- GMKMEZVLHJARHF-SYDPRGILSA-N meso-2,6-diaminopimelic acid Chemical compound [O-]C(=O)[C@@H]([NH3+])CCC[C@@H]([NH3+])C([O-])=O GMKMEZVLHJARHF-SYDPRGILSA-N 0.000 description 4
- 230000004048 modification Effects 0.000 description 4
- 238000012986 modification Methods 0.000 description 4
- 210000001616 monocyte Anatomy 0.000 description 4
- 238000010899 nucleation Methods 0.000 description 4
- HMFHBZSHGGEWLO-UHFFFAOYSA-N pentofuranose Chemical group OCC1OC(O)C(O)C1O HMFHBZSHGGEWLO-UHFFFAOYSA-N 0.000 description 4
- 108010073025 phenylalanylphenylalanine Proteins 0.000 description 4
- 230000026731 phosphorylation Effects 0.000 description 4
- 238000006366 phosphorylation reaction Methods 0.000 description 4
- 101150114864 plcA gene Proteins 0.000 description 4
- 229920000642 polymer Polymers 0.000 description 4
- 238000006116 polymerization reaction Methods 0.000 description 4
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 4
- 239000000047 product Substances 0.000 description 4
- 238000012797 qualification Methods 0.000 description 4
- 239000012925 reference material Substances 0.000 description 4
- 238000011160 research Methods 0.000 description 4
- 239000000725 suspension Substances 0.000 description 4
- 239000004474 valine Substances 0.000 description 4
- 229960004295 valine Drugs 0.000 description 4
- 206010069754 Acquired gene mutation Diseases 0.000 description 3
- YLTKNGYYPIWKHZ-ACZMJKKPSA-N Ala-Ala-Glu Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(O)=O YLTKNGYYPIWKHZ-ACZMJKKPSA-N 0.000 description 3
- JBVSSSZFNTXJDX-YTLHQDLWSA-N Ala-Ala-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](C)N JBVSSSZFNTXJDX-YTLHQDLWSA-N 0.000 description 3
- ZEXDYVGDZJBRMO-ACZMJKKPSA-N Ala-Asn-Gln Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N ZEXDYVGDZJBRMO-ACZMJKKPSA-N 0.000 description 3
- BLIMFWGRQKRCGT-YUMQZZPRSA-N Ala-Gly-Lys Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCCN BLIMFWGRQKRCGT-YUMQZZPRSA-N 0.000 description 3
- IFKQPMZRDQZSHI-GHCJXIJMSA-N Ala-Ile-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O IFKQPMZRDQZSHI-GHCJXIJMSA-N 0.000 description 3
- OYJCVIGKMXUVKB-GARJFASQSA-N Ala-Leu-Pro Chemical compound C[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N OYJCVIGKMXUVKB-GARJFASQSA-N 0.000 description 3
- MFMDKJIPHSWSBM-GUBZILKMSA-N Ala-Lys-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O MFMDKJIPHSWSBM-GUBZILKMSA-N 0.000 description 3
- QRIYOHQJRDHFKF-UWJYBYFXSA-N Ala-Tyr-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=C(O)C=C1 QRIYOHQJRDHFKF-UWJYBYFXSA-N 0.000 description 3
- VYSRNGOMGHOJCK-GUBZILKMSA-N Arg-Ala-Met Chemical compound C[C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N VYSRNGOMGHOJCK-GUBZILKMSA-N 0.000 description 3
- AWMAZIIEFPFHCP-RCWTZXSCSA-N Arg-Pro-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O AWMAZIIEFPFHCP-RCWTZXSCSA-N 0.000 description 3
- 241001167018 Aroa Species 0.000 description 3
- YRTOMUMWSTUQAX-FXQIFTODSA-N Asn-Pro-Asp Chemical compound NC(=O)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O YRTOMUMWSTUQAX-FXQIFTODSA-N 0.000 description 3
- JNNVNVRBYUJYGS-CIUDSAMLSA-N Asp-Leu-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O JNNVNVRBYUJYGS-CIUDSAMLSA-N 0.000 description 3
- HJCGDIGVVWETRO-ZPFDUUQYSA-N Asp-Lys-Ile Chemical compound CC[C@H](C)[C@H](NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CC(O)=O)C(O)=O HJCGDIGVVWETRO-ZPFDUUQYSA-N 0.000 description 3
- 108010078791 Carrier Proteins Proteins 0.000 description 3
- 101100491986 Emericella nidulans (strain FGSC A4 / ATCC 38163 / CBS 112.46 / NRRL 194 / M139) aromA gene Proteins 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- SBYVDRJAXWSXQL-AVGNSLFASA-N Glu-Asn-Phe Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O SBYVDRJAXWSXQL-AVGNSLFASA-N 0.000 description 3
- MUSGDMDGNGXULI-DCAQKATOSA-N Glu-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O MUSGDMDGNGXULI-DCAQKATOSA-N 0.000 description 3
- UZWUBBRJWFTHTD-LAEOZQHASA-N Glu-Val-Asn Chemical compound NC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCC(O)=O UZWUBBRJWFTHTD-LAEOZQHASA-N 0.000 description 3
- RJIVPOXLQFJRTG-LURJTMIESA-N Gly-Arg-Gly Chemical compound OC(=O)CNC(=O)[C@@H](NC(=O)CN)CCCN=C(N)N RJIVPOXLQFJRTG-LURJTMIESA-N 0.000 description 3
- VXKCPBPQEKKERH-IUCAKERBSA-N Gly-Arg-Pro Chemical compound NC(N)=NCCC[C@H](NC(=O)CN)C(=O)N1CCC[C@H]1C(O)=O VXKCPBPQEKKERH-IUCAKERBSA-N 0.000 description 3
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 3
- RVKIPWVMZANZLI-UHFFFAOYSA-N H-Lys-Trp-OH Natural products C1=CC=C2C(CC(NC(=O)C(N)CCCCN)C(O)=O)=CNC2=C1 RVKIPWVMZANZLI-UHFFFAOYSA-N 0.000 description 3
- 241000341655 Human papillomavirus type 16 Species 0.000 description 3
- DMSVBUWGDLYNLC-IAVJCBSLSA-N Ile-Ile-Phe Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 DMSVBUWGDLYNLC-IAVJCBSLSA-N 0.000 description 3
- UFRXVQGGPNSJRY-CYDGBPFRSA-N Ile-Met-Arg Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@H](C(O)=O)CCCN=C(N)N UFRXVQGGPNSJRY-CYDGBPFRSA-N 0.000 description 3
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 3
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 3
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 3
- OGCQGUIWMSBHRZ-CIUDSAMLSA-N Leu-Asn-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O OGCQGUIWMSBHRZ-CIUDSAMLSA-N 0.000 description 3
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 3
- 101100452614 Listeria monocytogenes serotype 1/2a (strain EGD / Mackaness) inlC gene Proteins 0.000 description 3
- 206010024641 Listeriosis Diseases 0.000 description 3
- SODXFJOPSCXOHE-IHRRRGAJSA-N Met-Leu-Leu Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O SODXFJOPSCXOHE-IHRRRGAJSA-N 0.000 description 3
- MSFSPUZXLOGKHJ-UHFFFAOYSA-N Muraminsaeure Natural products OC(=O)C(C)OC1C(N)C(O)OC(CO)C1O MSFSPUZXLOGKHJ-UHFFFAOYSA-N 0.000 description 3
- 108010087066 N2-tryptophyllysine Proteins 0.000 description 3
- 229910019142 PO4 Inorganic materials 0.000 description 3
- 108010013639 Peptidoglycan Proteins 0.000 description 3
- MCIXMYKSPQUMJG-SRVKXCTJSA-N Phe-Ser-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O MCIXMYKSPQUMJG-SRVKXCTJSA-N 0.000 description 3
- UUHXBJHVTVGSKM-BQBZGAKWSA-N Pro-Gly-Asn Chemical compound [H]N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O UUHXBJHVTVGSKM-BQBZGAKWSA-N 0.000 description 3
- QUBVFEANYYWBTM-VEVYYDQMSA-N Pro-Thr-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(O)=O QUBVFEANYYWBTM-VEVYYDQMSA-N 0.000 description 3
- FDMCIBSQRKFSTJ-RHYQMDGZSA-N Pro-Thr-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O FDMCIBSQRKFSTJ-RHYQMDGZSA-N 0.000 description 3
- 239000012980 RPMI-1640 medium Substances 0.000 description 3
- 108091081062 Repeated sequence (DNA) Proteins 0.000 description 3
- -1 Thr and Ala Chemical class 0.000 description 3
- LVHHEVGYAZGXDE-KDXUFGMBSA-N Thr-Ala-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](C)C(=O)N1CCC[C@@H]1C(=O)O)N)O LVHHEVGYAZGXDE-KDXUFGMBSA-N 0.000 description 3
- SGAOHNPSEPVAFP-ZDLURKLDSA-N Thr-Ser-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)NCC(O)=O SGAOHNPSEPVAFP-ZDLURKLDSA-N 0.000 description 3
- 108020004566 Transfer RNA Proteins 0.000 description 3
- KOVXHANYYYMBRF-IRIUXVKKSA-N Tyr-Glu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N)O KOVXHANYYYMBRF-IRIUXVKKSA-N 0.000 description 3
- 108090000848 Ubiquitin Proteins 0.000 description 3
- 102000044159 Ubiquitin Human genes 0.000 description 3
- ZLFHAAGHGQBQQN-AEJSXWLSSA-N Val-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](C(C)C)N ZLFHAAGHGQBQQN-AEJSXWLSSA-N 0.000 description 3
- ZLFHAAGHGQBQQN-GUBZILKMSA-N Val-Ala-Pro Natural products CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O ZLFHAAGHGQBQQN-GUBZILKMSA-N 0.000 description 3
- QPZMOUMNTGTEFR-ZKWXMUAHSA-N Val-Asn-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](C(C)C)N QPZMOUMNTGTEFR-ZKWXMUAHSA-N 0.000 description 3
- BMGOFDMKDVVGJG-NHCYSSNCSA-N Val-Asp-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N BMGOFDMKDVVGJG-NHCYSSNCSA-N 0.000 description 3
- XNLUVJPMPAZHCY-JYJNAYRXSA-N Val-Val-Phe Chemical compound CC(C)[C@H]([NH3+])C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C([O-])=O)CC1=CC=CC=C1 XNLUVJPMPAZHCY-JYJNAYRXSA-N 0.000 description 3
- 230000004913 activation Effects 0.000 description 3
- 230000001464 adherent effect Effects 0.000 description 3
- 108010013835 arginine glutamate Proteins 0.000 description 3
- 101150040872 aroE gene Proteins 0.000 description 3
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 3
- 230000001486 biosynthesis of amino acids Effects 0.000 description 3
- 230000015556 catabolic process Effects 0.000 description 3
- 230000006727 cell loss Effects 0.000 description 3
- 210000002421 cell wall Anatomy 0.000 description 3
- 235000012000 cholesterol Nutrition 0.000 description 3
- 230000007547 defect Effects 0.000 description 3
- 238000011156 evaluation Methods 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- 239000012595 freezing medium Substances 0.000 description 3
- 102000037865 fusion proteins Human genes 0.000 description 3
- 108020001507 fusion proteins Proteins 0.000 description 3
- 108010063718 gamma-glutamylaspartic acid Proteins 0.000 description 3
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 3
- 235000004554 glutamine Nutrition 0.000 description 3
- 230000002209 hydrophobic effect Effects 0.000 description 3
- 230000036039 immunity Effects 0.000 description 3
- 230000001939 inductive effect Effects 0.000 description 3
- 101150106970 inlA gene Proteins 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 239000010410 layer Substances 0.000 description 3
- 229960003136 leucine Drugs 0.000 description 3
- 108010057821 leucylproline Proteins 0.000 description 3
- 108010085203 methionylmethionine Proteins 0.000 description 3
- 231100000350 mutagenesis Toxicity 0.000 description 3
- 238000002703 mutagenesis Methods 0.000 description 3
- 230000000869 mutational effect Effects 0.000 description 3
- 230000006911 nucleation Effects 0.000 description 3
- 238000005457 optimization Methods 0.000 description 3
- 229910052760 oxygen Inorganic materials 0.000 description 3
- 239000001301 oxygen Substances 0.000 description 3
- 230000036961 partial effect Effects 0.000 description 3
- 230000001717 pathogenic effect Effects 0.000 description 3
- 239000008188 pellet Substances 0.000 description 3
- 108010024607 phenylalanylalanine Proteins 0.000 description 3
- 108010051242 phenylalanylserine Proteins 0.000 description 3
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 3
- 239000010452 phosphate Substances 0.000 description 3
- 239000002243 precursor Substances 0.000 description 3
- 238000001243 protein synthesis Methods 0.000 description 3
- 239000002904 solvent Substances 0.000 description 3
- 230000037439 somatic mutation Effects 0.000 description 3
- 238000004448 titration Methods 0.000 description 3
- 238000013519 translation Methods 0.000 description 3
- 230000005945 translocation Effects 0.000 description 3
- 238000010200 validation analysis Methods 0.000 description 3
- 239000013603 viral vector Substances 0.000 description 3
- MPUUQNGXJSEWTF-BYPYZUCNSA-N (S)-4-amino-5-oxopentanoic acid Chemical compound O=C[C@@H]([NH3+])CCC([O-])=O MPUUQNGXJSEWTF-BYPYZUCNSA-N 0.000 description 2
- KDCGOANMDULRCW-UHFFFAOYSA-N 7H-purine Chemical compound N1=CNC2=NC=NC2=C1 KDCGOANMDULRCW-UHFFFAOYSA-N 0.000 description 2
- ZGXJTSGNIOSYLO-UHFFFAOYSA-N 88755TAZ87 Chemical compound NCC(=O)CCC(O)=O ZGXJTSGNIOSYLO-UHFFFAOYSA-N 0.000 description 2
- DVWVZSJAYIJZFI-FXQIFTODSA-N Ala-Arg-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(O)=O DVWVZSJAYIJZFI-FXQIFTODSA-N 0.000 description 2
- SVBXIUDNTRTKHE-CIUDSAMLSA-N Ala-Arg-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=O SVBXIUDNTRTKHE-CIUDSAMLSA-N 0.000 description 2
- KXEVYGKATAMXJJ-ACZMJKKPSA-N Ala-Glu-Asp Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O KXEVYGKATAMXJJ-ACZMJKKPSA-N 0.000 description 2
- XYTNPQNAZREREP-XQXXSGGOSA-N Ala-Glu-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XYTNPQNAZREREP-XQXXSGGOSA-N 0.000 description 2
- RMAWDDRDTRSZIR-ZLUOBGJFSA-N Ala-Ser-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O RMAWDDRDTRSZIR-ZLUOBGJFSA-N 0.000 description 2
- LSMDIAAALJJLRO-XQXXSGGOSA-N Ala-Thr-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O LSMDIAAALJJLRO-XQXXSGGOSA-N 0.000 description 2
- ZXKNLCPUNZPFGY-LEWSCRJBSA-N Ala-Tyr-Pro Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N2CCC[C@@H]2C(=O)O)N ZXKNLCPUNZPFGY-LEWSCRJBSA-N 0.000 description 2
- CYXCAHZVPFREJD-LURJTMIESA-N Arg-Gly-Gly Chemical compound NC(=N)NCCC[C@H](N)C(=O)NCC(=O)NCC(O)=O CYXCAHZVPFREJD-LURJTMIESA-N 0.000 description 2
- CRCCTGPNZUCAHE-DCAQKATOSA-N Arg-His-Ser Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CO)C(O)=O)CC1=CN=CN1 CRCCTGPNZUCAHE-DCAQKATOSA-N 0.000 description 2
- XRNXPIGJPQHCPC-RCWTZXSCSA-N Arg-Thr-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CCCNC(N)=N)[C@@H](C)O)C(O)=O XRNXPIGJPQHCPC-RCWTZXSCSA-N 0.000 description 2
- HZPSDHRYYIORKR-WHFBIAKZSA-N Asn-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H](N)CC(N)=O HZPSDHRYYIORKR-WHFBIAKZSA-N 0.000 description 2
- XHFXZQHTLJVZBN-FXQIFTODSA-N Asn-Arg-Asn Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N)CN=C(N)N XHFXZQHTLJVZBN-FXQIFTODSA-N 0.000 description 2
- ZMWDUIIACVLIHK-GHCJXIJMSA-N Asn-Cys-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC(=O)N)N ZMWDUIIACVLIHK-GHCJXIJMSA-N 0.000 description 2
- QYXNFROWLZPWPC-FXQIFTODSA-N Asn-Glu-Gln Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O QYXNFROWLZPWPC-FXQIFTODSA-N 0.000 description 2
- MSBDSTRUMZFSEU-PEFMBERDSA-N Asn-Glu-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O MSBDSTRUMZFSEU-PEFMBERDSA-N 0.000 description 2
- JLNFZLNDHONLND-GARJFASQSA-N Asn-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(=O)N)N JLNFZLNDHONLND-GARJFASQSA-N 0.000 description 2
- LZLCLRQMUQWUHJ-GUBZILKMSA-N Asn-Lys-Gln Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N LZLCLRQMUQWUHJ-GUBZILKMSA-N 0.000 description 2
- NLDNNZKUSLAYFW-NHCYSSNCSA-N Asn-Lys-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O NLDNNZKUSLAYFW-NHCYSSNCSA-N 0.000 description 2
- GZXOUBTUAUAVHD-ACZMJKKPSA-N Asn-Ser-Glu Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O GZXOUBTUAUAVHD-ACZMJKKPSA-N 0.000 description 2
- MLJZMGIXXMTEPO-UBHSHLNASA-N Asn-Trp-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CO)C(O)=O MLJZMGIXXMTEPO-UBHSHLNASA-N 0.000 description 2
- HPNDBHLITCHRSO-WHFBIAKZSA-N Asp-Ala-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)NCC(O)=O HPNDBHLITCHRSO-WHFBIAKZSA-N 0.000 description 2
- UGIBTKGQVWFTGX-BIIVOSGPSA-N Asp-Asn-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC(=O)O)N)C(=O)O UGIBTKGQVWFTGX-BIIVOSGPSA-N 0.000 description 2
- JGDBHIVECJGXJA-FXQIFTODSA-N Asp-Asp-Arg Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O JGDBHIVECJGXJA-FXQIFTODSA-N 0.000 description 2
- WCFCYFDBMNFSPA-ACZMJKKPSA-N Asp-Asp-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCC(O)=O WCFCYFDBMNFSPA-ACZMJKKPSA-N 0.000 description 2
- DGKCOYGQLNWNCJ-ACZMJKKPSA-N Asp-Glu-Ser Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O DGKCOYGQLNWNCJ-ACZMJKKPSA-N 0.000 description 2
- SPWXXPFDTMYTRI-IUKAMOBKSA-N Asp-Ile-Thr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SPWXXPFDTMYTRI-IUKAMOBKSA-N 0.000 description 2
- USNJAPJZSGTTPX-XVSYOHENSA-N Asp-Phe-Thr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O USNJAPJZSGTTPX-XVSYOHENSA-N 0.000 description 2
- BRRPVTUFESPTCP-ACZMJKKPSA-N Asp-Ser-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O BRRPVTUFESPTCP-ACZMJKKPSA-N 0.000 description 2
- 108010003415 Aspartate Aminotransferases Proteins 0.000 description 2
- 102000004625 Aspartate Aminotransferases Human genes 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- 101100152417 Bacillus spizizenii (strain ATCC 23059 / NRRL B-14472 / W23) tarI gene Proteins 0.000 description 2
- 101100216993 Bacillus subtilis (strain 168) aroD gene Proteins 0.000 description 2
- 102100025570 Cancer/testis antigen 1 Human genes 0.000 description 2
- ALNKNYKSZPSLBD-ZDLURKLDSA-N Cys-Thr-Gly Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O ALNKNYKSZPSLBD-ZDLURKLDSA-N 0.000 description 2
- 101150029707 ERBB2 gene Proteins 0.000 description 2
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 2
- DRDSQGHKTLSNEA-GLLZPBPUSA-N Gln-Glu-Thr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O DRDSQGHKTLSNEA-GLLZPBPUSA-N 0.000 description 2
- NNXIQPMZGZUFJJ-AVGNSLFASA-N Gln-His-Lys Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)N)N NNXIQPMZGZUFJJ-AVGNSLFASA-N 0.000 description 2
- HWEINOMSWQSJDC-SRVKXCTJSA-N Gln-Leu-Arg Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O HWEINOMSWQSJDC-SRVKXCTJSA-N 0.000 description 2
- HPCOBEHVEHWREJ-DCAQKATOSA-N Gln-Lys-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O HPCOBEHVEHWREJ-DCAQKATOSA-N 0.000 description 2
- VNTGPISAOMAXRK-CIUDSAMLSA-N Gln-Pro-Ser Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O VNTGPISAOMAXRK-CIUDSAMLSA-N 0.000 description 2
- KUBFPYIMAGXGBT-ACZMJKKPSA-N Gln-Ser-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O KUBFPYIMAGXGBT-ACZMJKKPSA-N 0.000 description 2
- GLWXKFRTOHKGIT-ACZMJKKPSA-N Glu-Asn-Asn Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O GLWXKFRTOHKGIT-ACZMJKKPSA-N 0.000 description 2
- RJONUNZIMUXUOI-GUBZILKMSA-N Glu-Asn-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CCC(=O)O)N RJONUNZIMUXUOI-GUBZILKMSA-N 0.000 description 2
- VFZIDQZAEBORGY-GLLZPBPUSA-N Glu-Gln-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VFZIDQZAEBORGY-GLLZPBPUSA-N 0.000 description 2
- AUTNXSQEVVHSJK-YVNDNENWSA-N Glu-Glu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O AUTNXSQEVVHSJK-YVNDNENWSA-N 0.000 description 2
- VSRCAOIHMGCIJK-SRVKXCTJSA-N Glu-Leu-Arg Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O VSRCAOIHMGCIJK-SRVKXCTJSA-N 0.000 description 2
- VMKCPNBBPGGQBJ-GUBZILKMSA-N Glu-Leu-Asn Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCC(=O)O)N VMKCPNBBPGGQBJ-GUBZILKMSA-N 0.000 description 2
- OQXDUSZKISQQSS-GUBZILKMSA-N Glu-Lys-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O OQXDUSZKISQQSS-GUBZILKMSA-N 0.000 description 2
- MRWYPDWDZSLWJM-ACZMJKKPSA-N Glu-Ser-Asp Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O MRWYPDWDZSLWJM-ACZMJKKPSA-N 0.000 description 2
- GMVCSRBOSIUTFC-FXQIFTODSA-N Glu-Ser-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O GMVCSRBOSIUTFC-FXQIFTODSA-N 0.000 description 2
- XAXJIUAWAFVADB-VJBMBRPKSA-N Glu-Trp-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CNC4=CC=CC=C43)C(=O)O)NC(=O)[C@H](CCC(=O)O)N XAXJIUAWAFVADB-VJBMBRPKSA-N 0.000 description 2
- 108010070675 Glutathione transferase Proteins 0.000 description 2
- 102000005720 Glutathione transferase Human genes 0.000 description 2
- LJPIRKICOISLKN-WHFBIAKZSA-N Gly-Ala-Ser Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O LJPIRKICOISLKN-WHFBIAKZSA-N 0.000 description 2
- RPLLQZBOVIVGMX-QWRGUYRKSA-N Gly-Asp-Phe Chemical compound [H]NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O RPLLQZBOVIVGMX-QWRGUYRKSA-N 0.000 description 2
- TVTZEOHWHUVYCG-KYNKHSRBSA-N Gly-Thr-Thr Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O TVTZEOHWHUVYCG-KYNKHSRBSA-N 0.000 description 2
- JBCLFWXMTIKCCB-UHFFFAOYSA-N H-Gly-Phe-OH Natural products NCC(=O)NC(C(O)=O)CC1=CC=CC=C1 JBCLFWXMTIKCCB-UHFFFAOYSA-N 0.000 description 2
- 101000856237 Homo sapiens Cancer/testis antigen 1 Proteins 0.000 description 2
- QIHJTGSVGIPHIW-QSFUFRPTSA-N Ile-Asn-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](C(C)C)C(=O)O)N QIHJTGSVGIPHIW-QSFUFRPTSA-N 0.000 description 2
- PHIXPNQDGGILMP-YVNDNENWSA-N Ile-Glu-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N PHIXPNQDGGILMP-YVNDNENWSA-N 0.000 description 2
- AGGIYSLVUKVOPT-HTFCKZLJSA-N Ile-Ser-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)N AGGIYSLVUKVOPT-HTFCKZLJSA-N 0.000 description 2
- CNMOKANDJMLAIF-CIQUZCHMSA-N Ile-Thr-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O CNMOKANDJMLAIF-CIQUZCHMSA-N 0.000 description 2
- GVEODXUBBFDBPW-MGHWNKPDSA-N Ile-Tyr-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=C(O)C=C1 GVEODXUBBFDBPW-MGHWNKPDSA-N 0.000 description 2
- 206010062016 Immunosuppression Diseases 0.000 description 2
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- 241000194035 Lactococcus lactis Species 0.000 description 2
- PBCHMHROGNUXMK-DLOVCJGASA-N Leu-Ala-His Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CN=CN1 PBCHMHROGNUXMK-DLOVCJGASA-N 0.000 description 2
- HXWALXSAVBLTPK-NUTKFTJISA-N Leu-Ala-Trp Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CC(C)C)N HXWALXSAVBLTPK-NUTKFTJISA-N 0.000 description 2
- WGNOPSQMIQERPK-GARJFASQSA-N Leu-Asn-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N1CCC[C@@H]1C(=O)O)N WGNOPSQMIQERPK-GARJFASQSA-N 0.000 description 2
- WGNOPSQMIQERPK-UHFFFAOYSA-N Leu-Asn-Pro Natural products CC(C)CC(N)C(=O)NC(CC(=O)N)C(=O)N1CCCC1C(=O)O WGNOPSQMIQERPK-UHFFFAOYSA-N 0.000 description 2
- PVMPDMIKUVNOBD-CIUDSAMLSA-N Leu-Asp-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O PVMPDMIKUVNOBD-CIUDSAMLSA-N 0.000 description 2
- HRTRLSRYZZKPCO-BJDJZHNGSA-N Leu-Ile-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O HRTRLSRYZZKPCO-BJDJZHNGSA-N 0.000 description 2
- ZRHDPZAAWLXXIR-SRVKXCTJSA-N Leu-Lys-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O ZRHDPZAAWLXXIR-SRVKXCTJSA-N 0.000 description 2
- AXVIGSRGTMNSJU-YESZJQIVSA-N Leu-Tyr-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N2CCC[C@@H]2C(=O)O)N AXVIGSRGTMNSJU-YESZJQIVSA-N 0.000 description 2
- 108050006654 Lipocalin Proteins 0.000 description 2
- 102000019298 Lipocalin Human genes 0.000 description 2
- 241000186806 Listeria grayi Species 0.000 description 2
- 241000186814 Listeria welshimeri Species 0.000 description 2
- NFLFJGGKOHYZJF-BJDJZHNGSA-N Lys-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN NFLFJGGKOHYZJF-BJDJZHNGSA-N 0.000 description 2
- SJNZALDHDUYDBU-IHRRRGAJSA-N Lys-Arg-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(O)=O SJNZALDHDUYDBU-IHRRRGAJSA-N 0.000 description 2
- DNEJSAIMVANNPA-DCAQKATOSA-N Lys-Asn-Arg Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O DNEJSAIMVANNPA-DCAQKATOSA-N 0.000 description 2
- YFGWNAROEYWGNL-GUBZILKMSA-N Lys-Gln-Asn Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O YFGWNAROEYWGNL-GUBZILKMSA-N 0.000 description 2
- ZXEUFAVXODIPHC-GUBZILKMSA-N Lys-Glu-Asn Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ZXEUFAVXODIPHC-GUBZILKMSA-N 0.000 description 2
- PBIPLDMFHAICIP-DCAQKATOSA-N Lys-Glu-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O PBIPLDMFHAICIP-DCAQKATOSA-N 0.000 description 2
- JYXBNQOKPRQNQS-YTFOTSKYSA-N Lys-Ile-Ile Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JYXBNQOKPRQNQS-YTFOTSKYSA-N 0.000 description 2
- MYZMQWHPDAYKIE-SRVKXCTJSA-N Lys-Leu-Ala Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O MYZMQWHPDAYKIE-SRVKXCTJSA-N 0.000 description 2
- WBSCNDJQPKSPII-KKUMJFAQSA-N Lys-Lys-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O WBSCNDJQPKSPII-KKUMJFAQSA-N 0.000 description 2
- PLDJDCJLRCYPJB-VOAKCMCISA-N Lys-Lys-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O PLDJDCJLRCYPJB-VOAKCMCISA-N 0.000 description 2
- GHKXHCMRAUYLBS-CIUDSAMLSA-N Lys-Ser-Asn Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O GHKXHCMRAUYLBS-CIUDSAMLSA-N 0.000 description 2
- JMNRXRPBHFGXQX-GUBZILKMSA-N Lys-Ser-Glu Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O JMNRXRPBHFGXQX-GUBZILKMSA-N 0.000 description 2
- ZUGVARDEGWMMLK-SRVKXCTJSA-N Lys-Ser-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCCN ZUGVARDEGWMMLK-SRVKXCTJSA-N 0.000 description 2
- SQRLLZAQNOQCEG-KKUMJFAQSA-N Lys-Tyr-Ser Chemical compound NCCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CO)C(O)=O)CC1=CC=C(O)C=C1 SQRLLZAQNOQCEG-KKUMJFAQSA-N 0.000 description 2
- 102000043129 MHC class I family Human genes 0.000 description 2
- 108091054437 MHC class I family Proteins 0.000 description 2
- 101710175625 Maltose/maltodextrin-binding periplasmic protein Proteins 0.000 description 2
- 102000003939 Membrane transport proteins Human genes 0.000 description 2
- 108090000301 Membrane transport proteins Proteins 0.000 description 2
- MJVAVZPDRWSRRC-UHFFFAOYSA-N Menadione Chemical compound C1=CC=C2C(=O)C(C)=CC(=O)C2=C1 MJVAVZPDRWSRRC-UHFFFAOYSA-N 0.000 description 2
- ULNXMMYXQKGNPG-LPEHRKFASA-N Met-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCSC)N ULNXMMYXQKGNPG-LPEHRKFASA-N 0.000 description 2
- FSTWDRPCQQUJIT-NHCYSSNCSA-N Met-Val-Glu Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CCSC)N FSTWDRPCQQUJIT-NHCYSSNCSA-N 0.000 description 2
- RJQXTJLFIWVMTO-TYNCELHUSA-N Methicillin Chemical compound COC1=CC=CC(OC)=C1C(=O)N[C@@H]1C(=O)N2[C@@H](C(O)=O)C(C)(C)S[C@@H]21 RJQXTJLFIWVMTO-TYNCELHUSA-N 0.000 description 2
- 108010062010 N-Acetylmuramoyl-L-alanine Amidase Proteins 0.000 description 2
- 108010047562 NGR peptide Proteins 0.000 description 2
- 229930193140 Neomycin Natural products 0.000 description 2
- 108010038807 Oligopeptides Proteins 0.000 description 2
- 102000015636 Oligopeptides Human genes 0.000 description 2
- 229930182555 Penicillin Natural products 0.000 description 2
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 2
- OSBADCBXAMSPQD-YESZJQIVSA-N Phe-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CC=CC=C2)N OSBADCBXAMSPQD-YESZJQIVSA-N 0.000 description 2
- ZJPGOXWRFNKIQL-JYJNAYRXSA-N Phe-Pro-Pro Chemical compound C([C@H](N)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(O)=O)C1=CC=CC=C1 ZJPGOXWRFNKIQL-JYJNAYRXSA-N 0.000 description 2
- NMELOOXSGDRBRU-YUMQZZPRSA-N Pro-Glu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CCC(=O)O)NC(=O)[C@@H]1CCCN1 NMELOOXSGDRBRU-YUMQZZPRSA-N 0.000 description 2
- BRJGUPWVFXKBQI-XUXIUFHCSA-N Pro-Leu-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O BRJGUPWVFXKBQI-XUXIUFHCSA-N 0.000 description 2
- MRYUJHGPZQNOAD-IHRRRGAJSA-N Pro-Leu-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@@H]1CCCN1 MRYUJHGPZQNOAD-IHRRRGAJSA-N 0.000 description 2
- VWHJZETTZDAGOM-XUXIUFHCSA-N Pro-Lys-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O VWHJZETTZDAGOM-XUXIUFHCSA-N 0.000 description 2
- WOIFYRZPIORBRY-AVGNSLFASA-N Pro-Lys-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O WOIFYRZPIORBRY-AVGNSLFASA-N 0.000 description 2
- SWRNSCMUXRLHCR-ULQDDVLXSA-N Pro-Phe-Lys Chemical compound C([C@@H](C(=O)N[C@@H](CCCCN)C(O)=O)NC(=O)[C@H]1NCCC1)C1=CC=CC=C1 SWRNSCMUXRLHCR-ULQDDVLXSA-N 0.000 description 2
- RCYUBVHMVUHEBM-RCWTZXSCSA-N Pro-Pro-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O RCYUBVHMVUHEBM-RCWTZXSCSA-N 0.000 description 2
- LNICFEXCAHIJOR-DCAQKATOSA-N Pro-Ser-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O LNICFEXCAHIJOR-DCAQKATOSA-N 0.000 description 2
- IURWWZYKYPEANQ-HJGDQZAQSA-N Pro-Thr-Glu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O IURWWZYKYPEANQ-HJGDQZAQSA-N 0.000 description 2
- IIRBTQHFVNGPMQ-AVGNSLFASA-N Pro-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@@H]1CCCN1 IIRBTQHFVNGPMQ-AVGNSLFASA-N 0.000 description 2
- 239000012979 RPMI medium Substances 0.000 description 2
- 108090001066 Racemases and epimerases Proteins 0.000 description 2
- 102000004879 Racemases and epimerases Human genes 0.000 description 2
- 241000607142 Salmonella Species 0.000 description 2
- ZUGXSSFMTXKHJS-ZLUOBGJFSA-N Ser-Ala-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O ZUGXSSFMTXKHJS-ZLUOBGJFSA-N 0.000 description 2
- BYIROAKULFFTEK-CIUDSAMLSA-N Ser-Asp-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CO BYIROAKULFFTEK-CIUDSAMLSA-N 0.000 description 2
- LALNXSXEYFUUDD-GUBZILKMSA-N Ser-Glu-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O LALNXSXEYFUUDD-GUBZILKMSA-N 0.000 description 2
- UIPXCLNLUUAMJU-JBDRJPRFSA-N Ser-Ile-Ser Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O UIPXCLNLUUAMJU-JBDRJPRFSA-N 0.000 description 2
- QYSFWUIXDFJUDW-DCAQKATOSA-N Ser-Leu-Arg Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O QYSFWUIXDFJUDW-DCAQKATOSA-N 0.000 description 2
- NLOAIFSWUUFQFR-CIUDSAMLSA-N Ser-Leu-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O NLOAIFSWUUFQFR-CIUDSAMLSA-N 0.000 description 2
- UGGWCAFQPKANMW-FXQIFTODSA-N Ser-Met-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C)C(O)=O UGGWCAFQPKANMW-FXQIFTODSA-N 0.000 description 2
- NMZXJDSKEGFDLJ-DCAQKATOSA-N Ser-Pro-Lys Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CO)N)C(=O)N[C@@H](CCCCN)C(=O)O NMZXJDSKEGFDLJ-DCAQKATOSA-N 0.000 description 2
- DINQYZRMXGWWTG-GUBZILKMSA-N Ser-Pro-Pro Chemical compound OC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 DINQYZRMXGWWTG-GUBZILKMSA-N 0.000 description 2
- ILZAUMFXKSIUEF-SRVKXCTJSA-N Ser-Ser-Phe Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 ILZAUMFXKSIUEF-SRVKXCTJSA-N 0.000 description 2
- 241000264435 Streptococcus dysgalactiae subsp. equisimilis Species 0.000 description 2
- 235000014897 Streptococcus lactis Nutrition 0.000 description 2
- 241000193996 Streptococcus pyogenes Species 0.000 description 2
- 108700026226 TATA Box Proteins 0.000 description 2
- 239000004098 Tetracycline Substances 0.000 description 2
- 102000002933 Thioredoxin Human genes 0.000 description 2
- YOSLMIPKOUAHKI-OLHMAJIHSA-N Thr-Asp-Asp Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O YOSLMIPKOUAHKI-OLHMAJIHSA-N 0.000 description 2
- YBXMGKCLOPDEKA-NUMRIWBASA-N Thr-Asp-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O YBXMGKCLOPDEKA-NUMRIWBASA-N 0.000 description 2
- LGNBRHZANHMZHK-NUMRIWBASA-N Thr-Glu-Asp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N)O LGNBRHZANHMZHK-NUMRIWBASA-N 0.000 description 2
- XOTBWOCSLMBGMF-SUSMZKCASA-N Thr-Glu-Thr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XOTBWOCSLMBGMF-SUSMZKCASA-N 0.000 description 2
- MGJLBZFUXUGMML-VOAKCMCISA-N Thr-Lys-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N)O MGJLBZFUXUGMML-VOAKCMCISA-N 0.000 description 2
- LKJCABTUFGTPPY-HJGDQZAQSA-N Thr-Pro-Gln Chemical compound C[C@@H](O)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O LKJCABTUFGTPPY-HJGDQZAQSA-N 0.000 description 2
- WPSKTVVMQCXPRO-BWBBJGPYSA-N Thr-Ser-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O WPSKTVVMQCXPRO-BWBBJGPYSA-N 0.000 description 2
- VBMOVTMNHWPZJR-SUSMZKCASA-N Thr-Thr-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O VBMOVTMNHWPZJR-SUSMZKCASA-N 0.000 description 2
- CSNBWOJOEOPYIJ-UVOCVTCTSA-N Thr-Thr-Lys Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(O)=O CSNBWOJOEOPYIJ-UVOCVTCTSA-N 0.000 description 2
- COYHRQWNJDJCNA-NUJDXYNKSA-N Thr-Thr-Thr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O COYHRQWNJDJCNA-NUJDXYNKSA-N 0.000 description 2
- 102000003929 Transaminases Human genes 0.000 description 2
- 108090000340 Transaminases Proteins 0.000 description 2
- GLNADSQYFUSGOU-GPTZEZBUSA-J Trypan blue Chemical compound [Na+].[Na+].[Na+].[Na+].C1=C(S([O-])(=O)=O)C=C2C=C(S([O-])(=O)=O)C(/N=N/C3=CC=C(C=C3C)C=3C=C(C(=CC=3)\N=N\C=3C(=CC4=CC(=CC(N)=C4C=3O)S([O-])(=O)=O)S([O-])(=O)=O)C)=C(O)C2=C1N GLNADSQYFUSGOU-GPTZEZBUSA-J 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 2
- OTJMMKPMLUNTQT-AVGNSLFASA-N Val-Leu-Arg Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](C(C)C)N OTJMMKPMLUNTQT-AVGNSLFASA-N 0.000 description 2
- NZGOVKLVQNOEKP-YDHLFZDLSA-N Val-Phe-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(=O)N)C(=O)O)N NZGOVKLVQNOEKP-YDHLFZDLSA-N 0.000 description 2
- PZTZYZUTCPZWJH-FXQIFTODSA-N Val-Ser-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)O)N PZTZYZUTCPZWJH-FXQIFTODSA-N 0.000 description 2
- 230000002378 acidificating effect Effects 0.000 description 2
- 108010008685 alanyl-glutamyl-aspartic acid Proteins 0.000 description 2
- 108010012597 alveolysin Proteins 0.000 description 2
- 229960000723 ampicillin Drugs 0.000 description 2
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 2
- 230000003321 amplification Effects 0.000 description 2
- 230000003698 anagen phase Effects 0.000 description 2
- 238000000540 analysis of variance Methods 0.000 description 2
- 101150029940 argJ gene Proteins 0.000 description 2
- 101150090235 aroB gene Proteins 0.000 description 2
- 101150042732 aroC gene Proteins 0.000 description 2
- 101150102858 aroD gene Proteins 0.000 description 2
- 101150108612 aroQ gene Proteins 0.000 description 2
- 235000003704 aspartic acid Nutrition 0.000 description 2
- 108010069205 aspartyl-phenylalanine Proteins 0.000 description 2
- 108010093581 aspartyl-proline Proteins 0.000 description 2
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 2
- 230000036952 cancer formation Effects 0.000 description 2
- 238000002619 cancer immunotherapy Methods 0.000 description 2
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 2
- 230000032823 cell division Effects 0.000 description 2
- 230000010261 cell growth Effects 0.000 description 2
- 239000013592 cell lysate Substances 0.000 description 2
- 230000003833 cell viability Effects 0.000 description 2
- 238000004587 chromatography analysis Methods 0.000 description 2
- 230000002759 chromosomal effect Effects 0.000 description 2
- 238000010367 cloning Methods 0.000 description 2
- 230000000295 complement effect Effects 0.000 description 2
- 230000021615 conjugation Effects 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- 238000009792 diffusion process Methods 0.000 description 2
- 239000003814 drug Substances 0.000 description 2
- 229940079593 drug Drugs 0.000 description 2
- 239000002158 endotoxin Substances 0.000 description 2
- 230000007613 environmental effect Effects 0.000 description 2
- 229960003276 erythromycin Drugs 0.000 description 2
- 238000002181 esophagogastroduodenoscopy Methods 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 230000008014 freezing Effects 0.000 description 2
- 238000007710 freezing Methods 0.000 description 2
- 239000007789 gas Substances 0.000 description 2
- 238000012224 gene deletion Methods 0.000 description 2
- 210000002980 germ line cell Anatomy 0.000 description 2
- 230000006801 homologous recombination Effects 0.000 description 2
- 238000002744 homologous recombination Methods 0.000 description 2
- 230000002779 inactivation Effects 0.000 description 2
- 230000036512 infertility Effects 0.000 description 2
- 239000012499 inoculation medium Substances 0.000 description 2
- 101150014059 ispD gene Proteins 0.000 description 2
- 101150022203 ispDF gene Proteins 0.000 description 2
- 229960000318 kanamycin Drugs 0.000 description 2
- 229930027917 kanamycin Natural products 0.000 description 2
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 2
- 229930182823 kanamycin A Natural products 0.000 description 2
- 101150087199 leuA gene Proteins 0.000 description 2
- 229920006008 lipopolysaccharide Polymers 0.000 description 2
- 210000003712 lysosome Anatomy 0.000 description 2
- 230000001868 lysosomic effect Effects 0.000 description 2
- 229920002521 macromolecule Polymers 0.000 description 2
- 210000004962 mammalian cell Anatomy 0.000 description 2
- 229930182817 methionine Natural products 0.000 description 2
- 229960003085 meticillin Drugs 0.000 description 2
- 229960004927 neomycin Drugs 0.000 description 2
- 238000003199 nucleic acid amplification method Methods 0.000 description 2
- 244000052769 pathogen Species 0.000 description 2
- 230000007918 pathogenicity Effects 0.000 description 2
- 229940049954 penicillin Drugs 0.000 description 2
- 229910052698 phosphorus Inorganic materials 0.000 description 2
- 230000004260 plant-type cell wall biogenesis Effects 0.000 description 2
- 230000002028 premature Effects 0.000 description 2
- 108010031719 prolyl-serine Proteins 0.000 description 2
- 108010049718 pseudouridine synthases Proteins 0.000 description 2
- 230000008707 rearrangement Effects 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 238000000611 regression analysis Methods 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 230000003248 secreting effect Effects 0.000 description 2
- 108010048397 seryl-lysyl-leucine Proteins 0.000 description 2
- 230000037432 silent mutation Effects 0.000 description 2
- 239000000243 solution Substances 0.000 description 2
- 210000001082 somatic cell Anatomy 0.000 description 2
- 230000000392 somatic effect Effects 0.000 description 2
- ATHGHQPFGPMSJY-UHFFFAOYSA-N spermidine Chemical compound NCCCCNCCCN ATHGHQPFGPMSJY-UHFFFAOYSA-N 0.000 description 2
- 230000000638 stimulation Effects 0.000 description 2
- 239000011550 stock solution Substances 0.000 description 2
- 108010075210 streptolysin O Proteins 0.000 description 2
- 238000010998 test method Methods 0.000 description 2
- 229960002180 tetracycline Drugs 0.000 description 2
- 229930101283 tetracycline Natural products 0.000 description 2
- 235000019364 tetracycline Nutrition 0.000 description 2
- 150000003522 tetracyclines Chemical class 0.000 description 2
- 108060008226 thioredoxin Proteins 0.000 description 2
- 229940094937 thioredoxin Drugs 0.000 description 2
- 108010071097 threonyl-lysyl-proline Proteins 0.000 description 2
- 230000009466 transformation Effects 0.000 description 2
- 230000001131 transforming effect Effects 0.000 description 2
- 101150059846 trpS gene Proteins 0.000 description 2
- 230000007923 virulence factor Effects 0.000 description 2
- 239000000304 virulence factor Substances 0.000 description 2
- 235000013343 vitamin Nutrition 0.000 description 2
- 239000011782 vitamin Substances 0.000 description 2
- 229940088594 vitamin Drugs 0.000 description 2
- 229930003231 vitamin Natural products 0.000 description 2
- HKZAAJSTFUZYTO-LURJTMIESA-N (2s)-2-[[2-[[2-[[2-[(2-aminoacetyl)amino]acetyl]amino]acetyl]amino]acetyl]amino]-3-hydroxypropanoic acid Chemical compound NCC(=O)NCC(=O)NCC(=O)NCC(=O)N[C@@H](CO)C(O)=O HKZAAJSTFUZYTO-LURJTMIESA-N 0.000 description 1
- KWIBRHGQCHIETC-CWSFPBCASA-N (7s)-3-[(3s)-3-[[(2r)-2-acetamido-4-[(2r,3s,4r,5r,6s)-5-amino-2-[[[[(2r,3s,4r)-5-(2,4-dioxopyrimidin-1-yl)-3,4-dihydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-hydroxyphosphoryl]oxymethyl]-3,6-dihydroxyoxan-4-yl]oxy-2-methyl-3-oxopentanoyl]amino]-4-[[(2 Chemical compound O[C@H]1[C@H](OC(C)C(=O)[C@@](C)(NC(C)=O)C(=O)N[C@@H](CCC(C(N)C(O)=O)C(=O)CC[C@H](N)C(O)=O)C(=O)N[C@H](C)C(=O)N[C@H](C)C(O)=O)[C@@H](N)[C@@H](O)O[C@@H]1COP(O)(=O)OP(O)(=O)OC[C@@H]1[C@@H](O)[C@@H](O)C(N2C(NC(=O)C=C2)=O)O1 KWIBRHGQCHIETC-CWSFPBCASA-N 0.000 description 1
- GHOKWGTUZJEAQD-ZETCQYMHSA-N (D)-(+)-Pantothenic acid Chemical compound OCC(C)(C)[C@@H](O)C(=O)NCCC(O)=O GHOKWGTUZJEAQD-ZETCQYMHSA-N 0.000 description 1
- NQEQTYPJSIEPHW-UHFFFAOYSA-N 1-C-(indol-3-yl)glycerol 3-phosphate Chemical compound C1=CC=C2C(C(O)C(COP(O)(O)=O)O)=CNC2=C1 NQEQTYPJSIEPHW-UHFFFAOYSA-N 0.000 description 1
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 1
- ZIIUUSVHCHPIQD-UHFFFAOYSA-N 2,4,6-trimethyl-N-[3-(trifluoromethyl)phenyl]benzenesulfonamide Chemical compound CC1=CC(C)=CC(C)=C1S(=O)(=O)NC1=CC=CC(C(F)(F)F)=C1 ZIIUUSVHCHPIQD-UHFFFAOYSA-N 0.000 description 1
- QMOQBVOBWVNSNO-UHFFFAOYSA-N 2-[[2-[[2-[(2-azaniumylacetyl)amino]acetyl]amino]acetyl]amino]acetate Chemical compound NCC(=O)NCC(=O)NCC(=O)NCC(O)=O QMOQBVOBWVNSNO-UHFFFAOYSA-N 0.000 description 1
- 108010030844 2-methylcitrate synthase Proteins 0.000 description 1
- KPGXRSRHYNQIFN-UHFFFAOYSA-N 2-oxoglutaric acid Chemical compound OC(=O)CCC(=O)C(O)=O KPGXRSRHYNQIFN-UHFFFAOYSA-N 0.000 description 1
- KDVFRMMRZOCFLS-UHFFFAOYSA-N 2-oxopentanoic acid Chemical compound CCCC(=O)C(O)=O KDVFRMMRZOCFLS-UHFFFAOYSA-N 0.000 description 1
- WVMWZWGZRAXUBK-SYTVJDICSA-M 3-dehydroquinate Chemical compound O[C@@H]1C[C@](O)(C([O-])=O)CC(=O)[C@H]1O WVMWZWGZRAXUBK-SYTVJDICSA-M 0.000 description 1
- WVMWZWGZRAXUBK-UHFFFAOYSA-N 3-dehydroquinic acid Natural products OC1CC(O)(C(O)=O)CC(=O)C1O WVMWZWGZRAXUBK-UHFFFAOYSA-N 0.000 description 1
- KQPKMEYBZUPZGK-UHFFFAOYSA-N 4-[(4-azido-2-nitroanilino)methyl]-5-(hydroxymethyl)-2-methylpyridin-3-ol Chemical compound CC1=NC=C(CO)C(CNC=2C(=CC(=CC=2)N=[N+]=[N-])[N+]([O-])=O)=C1O KQPKMEYBZUPZGK-UHFFFAOYSA-N 0.000 description 1
- 108010075604 5-Methyltetrahydrofolate-Homocysteine S-Methyltransferase Proteins 0.000 description 1
- 102000011848 5-Methyltetrahydrofolate-Homocysteine S-Methyltransferase Human genes 0.000 description 1
- GOZMBJCYMQQACI-UHFFFAOYSA-N 6,7-dimethyl-3-[[methyl-[2-[methyl-[[1-[3-(trifluoromethyl)phenyl]indol-3-yl]methyl]amino]ethyl]amino]methyl]chromen-4-one;dihydrochloride Chemical compound Cl.Cl.C=1OC2=CC(C)=C(C)C=C2C(=O)C=1CN(C)CCN(C)CC(C1=CC=CC=C11)=CN1C1=CC=CC(C(F)(F)F)=C1 GOZMBJCYMQQACI-UHFFFAOYSA-N 0.000 description 1
- 108700014229 ABC-type teichoic acid transporter activity proteins Proteins 0.000 description 1
- RZVAJINKPMORJF-UHFFFAOYSA-N Acetaminophen Chemical compound CC(=O)NC1=CC=C(O)C=C1 RZVAJINKPMORJF-UHFFFAOYSA-N 0.000 description 1
- 206010000871 Acute monocytic leukaemia Diseases 0.000 description 1
- BYXHQQCXAJARLQ-ZLUOBGJFSA-N Ala-Ala-Ala Chemical group C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O BYXHQQCXAJARLQ-ZLUOBGJFSA-N 0.000 description 1
- TTXMOJWKNRJWQJ-FXQIFTODSA-N Ala-Arg-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CCCN=C(N)N TTXMOJWKNRJWQJ-FXQIFTODSA-N 0.000 description 1
- LBJYAILUMSUTAM-ZLUOBGJFSA-N Ala-Asn-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O LBJYAILUMSUTAM-ZLUOBGJFSA-N 0.000 description 1
- YBPLKDWJFYCZSV-ZLUOBGJFSA-N Ala-Asn-Cys Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CS)C(=O)O)N YBPLKDWJFYCZSV-ZLUOBGJFSA-N 0.000 description 1
- CVGNCMIULZNYES-WHFBIAKZSA-N Ala-Asn-Gly Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O CVGNCMIULZNYES-WHFBIAKZSA-N 0.000 description 1
- NINQYGGNRIBFSC-CIUDSAMLSA-N Ala-Lys-Ser Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CO)C(O)=O NINQYGGNRIBFSC-CIUDSAMLSA-N 0.000 description 1
- RUXQNKVQSKOOBS-JURCDPSOSA-N Ala-Phe-Ile Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O RUXQNKVQSKOOBS-JURCDPSOSA-N 0.000 description 1
- MSWSRLGNLKHDEI-ACZMJKKPSA-N Ala-Ser-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O MSWSRLGNLKHDEI-ACZMJKKPSA-N 0.000 description 1
- QOIGKCBMXUCDQU-KDXUFGMBSA-N Ala-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](C)N)O QOIGKCBMXUCDQU-KDXUFGMBSA-N 0.000 description 1
- 101100163490 Alkalihalobacillus halodurans (strain ATCC BAA-125 / DSM 18197 / FERM 7344 / JCM 9153 / C-125) aroA1 gene Proteins 0.000 description 1
- 108700028369 Alleles Proteins 0.000 description 1
- 241000388169 Alphapapillomavirus 7 Species 0.000 description 1
- 102000052866 Amino Acyl-tRNA Synthetases Human genes 0.000 description 1
- 108700028939 Amino Acyl-tRNA Synthetases Proteins 0.000 description 1
- QGZKDVFQNNGYKY-UHFFFAOYSA-O Ammonium Chemical compound [NH4+] QGZKDVFQNNGYKY-UHFFFAOYSA-O 0.000 description 1
- 241000024188 Andala Species 0.000 description 1
- 108010037870 Anthranilate Synthase Proteins 0.000 description 1
- 208000007860 Anus Neoplasms Diseases 0.000 description 1
- 101000640990 Arabidopsis thaliana Tryptophan-tRNA ligase, chloroplastic/mitochondrial Proteins 0.000 description 1
- SQKPKIJVWHAWNF-DCAQKATOSA-N Arg-Asp-Lys Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(O)=O SQKPKIJVWHAWNF-DCAQKATOSA-N 0.000 description 1
- ISJWBVIYRBAXEB-CIUDSAMLSA-N Arg-Ser-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O ISJWBVIYRBAXEB-CIUDSAMLSA-N 0.000 description 1
- SWLOHUMCUDRTCL-ZLUOBGJFSA-N Asn-Ala-Asn Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N SWLOHUMCUDRTCL-ZLUOBGJFSA-N 0.000 description 1
- LXTGAOAXPSJWOU-DCAQKATOSA-N Asn-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(=O)N)N LXTGAOAXPSJWOU-DCAQKATOSA-N 0.000 description 1
- HUZGPXBILPMCHM-IHRRRGAJSA-N Asn-Arg-Phe Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O HUZGPXBILPMCHM-IHRRRGAJSA-N 0.000 description 1
- GMUOCGCDOYYWPD-FXQIFTODSA-N Asn-Pro-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O GMUOCGCDOYYWPD-FXQIFTODSA-N 0.000 description 1
- QYRMBFWDSFGSFC-OLHMAJIHSA-N Asn-Thr-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N)O QYRMBFWDSFGSFC-OLHMAJIHSA-N 0.000 description 1
- FMNBYVSGRCXWEK-FOHZUACHSA-N Asn-Thr-Gly Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O FMNBYVSGRCXWEK-FOHZUACHSA-N 0.000 description 1
- CELPEWWLSXMVPH-CIUDSAMLSA-N Asp-Asp-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC(O)=O CELPEWWLSXMVPH-CIUDSAMLSA-N 0.000 description 1
- OGTCOKZFOJIZFG-CIUDSAMLSA-N Asp-His-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(O)=O)C(O)=O OGTCOKZFOJIZFG-CIUDSAMLSA-N 0.000 description 1
- TZOZNVLBTAFJRW-UGYAYLCHSA-N Asp-Ile-Asp Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CC(=O)O)N TZOZNVLBTAFJRW-UGYAYLCHSA-N 0.000 description 1
- YFSLJHLQOALGSY-ZPFDUUQYSA-N Asp-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)O)N YFSLJHLQOALGSY-ZPFDUUQYSA-N 0.000 description 1
- BJDHEININLSZOT-KKUMJFAQSA-N Asp-Tyr-Lys Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCCN)C(O)=O BJDHEININLSZOT-KKUMJFAQSA-N 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 108020004652 Aspartate-Semialdehyde Dehydrogenase Proteins 0.000 description 1
- 238000011725 BALB/c mouse Methods 0.000 description 1
- 241000193830 Bacillus <bacterium> Species 0.000 description 1
- 241000193738 Bacillus anthracis Species 0.000 description 1
- 101100012355 Bacillus anthracis fabH1 gene Proteins 0.000 description 1
- 101100427060 Bacillus spizizenii (strain ATCC 23059 / NRRL B-14472 / W23) thyA1 gene Proteins 0.000 description 1
- 235000014469 Bacillus subtilis Nutrition 0.000 description 1
- 101100072559 Bacillus subtilis (strain 168) alsS gene Proteins 0.000 description 1
- 101100221537 Bacillus subtilis (strain 168) comK gene Proteins 0.000 description 1
- 101100012357 Bacillus subtilis (strain 168) fabHA gene Proteins 0.000 description 1
- 101100394745 Bacillus subtilis (strain 168) hepT gene Proteins 0.000 description 1
- 241000193388 Bacillus thuringiensis Species 0.000 description 1
- 241000212384 Bifora Species 0.000 description 1
- 101100378273 Brachyspira hyodysenteriae acpP gene Proteins 0.000 description 1
- 102100032912 CD44 antigen Human genes 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- UXVMQQNJUSDDNG-UHFFFAOYSA-L Calcium chloride Chemical compound [Cl-].[Cl-].[Ca+2] UXVMQQNJUSDDNG-UHFFFAOYSA-L 0.000 description 1
- 101100339117 Campylobacter jejuni subsp. jejuni serotype O:2 (strain ATCC 700819 / NCTC 11168) hisF1 gene Proteins 0.000 description 1
- 102000005367 Carboxypeptidases Human genes 0.000 description 1
- 108010006303 Carboxypeptidases Proteins 0.000 description 1
- 208000005623 Carcinogenesis Diseases 0.000 description 1
- 206010008342 Cervix carcinoma Diseases 0.000 description 1
- 229920002101 Chitin Polymers 0.000 description 1
- 101100227595 Chlamydia pneumoniae folKP gene Proteins 0.000 description 1
- 108020004638 Circular DNA Proteins 0.000 description 1
- 108091062157 Cis-regulatory element Proteins 0.000 description 1
- 108010071536 Citrate (Si)-synthase Proteins 0.000 description 1
- 102000006732 Citrate synthase Human genes 0.000 description 1
- 101100163308 Clostridium perfringens (strain 13 / Type A) argR1 gene Proteins 0.000 description 1
- 101100538459 Clostridium perfringens (strain 13 / Type A) truA1 gene Proteins 0.000 description 1
- 101100100749 Clostridium perfringens (strain 13 / Type A) truA2 gene Proteins 0.000 description 1
- 101710170530 Cysteine synthase A Proteins 0.000 description 1
- 108090000695 Cytokines Proteins 0.000 description 1
- 102000004127 Cytokines Human genes 0.000 description 1
- 108010078895 D-Alanine Transaminase Proteins 0.000 description 1
- 239000003298 DNA probe Substances 0.000 description 1
- 101100465553 Dictyostelium discoideum psmB6 gene Proteins 0.000 description 1
- 102100025137 Early activation antigen CD69 Human genes 0.000 description 1
- 101100507308 Enterococcus faecalis mvaS gene Proteins 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 101100337176 Escherichia coli (strain K12) gltB gene Proteins 0.000 description 1
- 101100505027 Escherichia coli (strain K12) gltD gene Proteins 0.000 description 1
- 101100155531 Escherichia coli (strain K12) ispU gene Proteins 0.000 description 1
- 101100139916 Escherichia coli (strain K12) rarA gene Proteins 0.000 description 1
- 101100540759 Escherichia coli (strain K12) wecE gene Proteins 0.000 description 1
- 101100153154 Escherichia phage T5 thy gene Proteins 0.000 description 1
- 241000589565 Flavobacterium Species 0.000 description 1
- 108050000897 Formate/nitrite transporters Proteins 0.000 description 1
- 101710140958 Formimidoyltetrahydrofolate cyclodeaminase Proteins 0.000 description 1
- 108090000926 GMP synthase (glutamine-hydrolyzing) Proteins 0.000 description 1
- 102100033452 GMP synthase [glutamine-hydrolyzing] Human genes 0.000 description 1
- XZLLTYBONVKGLO-SDDRHHMPSA-N Gln-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(=O)N)N)C(=O)O XZLLTYBONVKGLO-SDDRHHMPSA-N 0.000 description 1
- WOMUDRVDJMHTCV-DCAQKATOSA-N Glu-Arg-Arg Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O WOMUDRVDJMHTCV-DCAQKATOSA-N 0.000 description 1
- WLIPTFCZLHCNFD-LPEHRKFASA-N Glu-Gln-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCC(=O)O)N)C(=O)O WLIPTFCZLHCNFD-LPEHRKFASA-N 0.000 description 1
- BUZMZDDKFCSKOT-CIUDSAMLSA-N Glu-Glu-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O BUZMZDDKFCSKOT-CIUDSAMLSA-N 0.000 description 1
- KASDBWKLWJKTLJ-GUBZILKMSA-N Glu-Glu-Met Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCSC)C(O)=O KASDBWKLWJKTLJ-GUBZILKMSA-N 0.000 description 1
- LGYCLOCORAEQSZ-PEFMBERDSA-N Glu-Ile-Asp Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O LGYCLOCORAEQSZ-PEFMBERDSA-N 0.000 description 1
- XTZDZAXYPDISRR-MNXVOIDGSA-N Glu-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N XTZDZAXYPDISRR-MNXVOIDGSA-N 0.000 description 1
- UGSVSNXPJJDJKL-SDDRHHMPSA-N Glu-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)O)N UGSVSNXPJJDJKL-SDDRHHMPSA-N 0.000 description 1
- UJMNFCAHLYKWOZ-DCAQKATOSA-N Glu-Lys-Gln Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(O)=O UJMNFCAHLYKWOZ-DCAQKATOSA-N 0.000 description 1
- HRBYTAIBKPNZKQ-AVGNSLFASA-N Glu-Lys-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCC(O)=O HRBYTAIBKPNZKQ-AVGNSLFASA-N 0.000 description 1
- SOEPMWQCTJITPZ-SRVKXCTJSA-N Glu-Met-Lys Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N SOEPMWQCTJITPZ-SRVKXCTJSA-N 0.000 description 1
- CBWKURKPYSLMJV-SOUVJXGZSA-N Glu-Phe-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=CC=C2)NC(=O)[C@H](CCC(=O)O)N)C(=O)O CBWKURKPYSLMJV-SOUVJXGZSA-N 0.000 description 1
- FGSGPLRPQCZBSQ-AVGNSLFASA-N Glu-Phe-Ser Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O FGSGPLRPQCZBSQ-AVGNSLFASA-N 0.000 description 1
- IDEODOAVGCMUQV-GUBZILKMSA-N Glu-Ser-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O IDEODOAVGCMUQV-GUBZILKMSA-N 0.000 description 1
- HMJULNMJWOZNFI-XHNCKOQMSA-N Glu-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)[C@H](CCC(=O)O)N)C(=O)O HMJULNMJWOZNFI-XHNCKOQMSA-N 0.000 description 1
- DMYACXMQUABZIQ-NRPADANISA-N Glu-Ser-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O DMYACXMQUABZIQ-NRPADANISA-N 0.000 description 1
- CAQXJMUDOLSBPF-SUSMZKCASA-N Glu-Thr-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O CAQXJMUDOLSBPF-SUSMZKCASA-N 0.000 description 1
- GRIRDMVMJJDZKV-RCOVLWMOSA-N Gly-Asn-Val Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O GRIRDMVMJJDZKV-RCOVLWMOSA-N 0.000 description 1
- FZQLXNIMCPJVJE-YUMQZZPRSA-N Gly-Asp-Leu Chemical compound [H]NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O FZQLXNIMCPJVJE-YUMQZZPRSA-N 0.000 description 1
- QGZSAHIZRQHCEQ-QWRGUYRKSA-N Gly-Asp-Tyr Chemical compound NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 QGZSAHIZRQHCEQ-QWRGUYRKSA-N 0.000 description 1
- MHXKHKWHPNETGG-QWRGUYRKSA-N Gly-Lys-Leu Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O MHXKHKWHPNETGG-QWRGUYRKSA-N 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 108010001483 Glycogen Synthase Proteins 0.000 description 1
- 108010006464 Hemolysin Proteins Proteins 0.000 description 1
- KZTLOHBDLMIFSH-XVYDVKMFSA-N His-Ala-Asp Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(O)=O KZTLOHBDLMIFSH-XVYDVKMFSA-N 0.000 description 1
- FHKZHRMERJUXRJ-DCAQKATOSA-N His-Ser-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC1=CN=CN1 FHKZHRMERJUXRJ-DCAQKATOSA-N 0.000 description 1
- NBWATNYAUVSAEQ-ZEILLAHLSA-N His-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CC1=CN=CN1)N)O NBWATNYAUVSAEQ-ZEILLAHLSA-N 0.000 description 1
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 1
- 101000934374 Homo sapiens Early activation antigen CD69 Proteins 0.000 description 1
- 101001057504 Homo sapiens Interferon-stimulated gene 20 kDa protein Proteins 0.000 description 1
- 101001055144 Homo sapiens Interleukin-2 receptor subunit alpha Proteins 0.000 description 1
- 101001111984 Homo sapiens N-acylneuraminate-9-phosphatase Proteins 0.000 description 1
- 102000004157 Hydrolases Human genes 0.000 description 1
- 108090000604 Hydrolases Proteins 0.000 description 1
- 108010042653 IgA receptor Proteins 0.000 description 1
- DFJJAVZIHDFOGQ-MNXVOIDGSA-N Ile-Glu-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N DFJJAVZIHDFOGQ-MNXVOIDGSA-N 0.000 description 1
- SVBAHOMTJRFSIC-SXTJYALSSA-N Ile-Ile-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(=O)N)C(=O)O)N SVBAHOMTJRFSIC-SXTJYALSSA-N 0.000 description 1
- KTNGVMMGIQWIDV-OSUNSFLBSA-N Ile-Pro-Thr Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O KTNGVMMGIQWIDV-OSUNSFLBSA-N 0.000 description 1
- HJDZMPFEXINXLO-QPHKQPEJSA-N Ile-Thr-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)N HJDZMPFEXINXLO-QPHKQPEJSA-N 0.000 description 1
- BCISUQVFDGYZBO-QSFUFRPTSA-N Ile-Val-Asp Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(O)=O BCISUQVFDGYZBO-QSFUFRPTSA-N 0.000 description 1
- 108010065920 Insulin Lispro Proteins 0.000 description 1
- 108010002352 Interleukin-1 Proteins 0.000 description 1
- 108010002350 Interleukin-2 Proteins 0.000 description 1
- 102100026878 Interleukin-2 receptor subunit alpha Human genes 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 229930195714 L-glutamate Natural products 0.000 description 1
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 1
- 229930182821 L-proline Natural products 0.000 description 1
- 125000000174 L-prolyl group Chemical group [H]N1C([H])([H])C([H])([H])C([H])([H])[C@@]1([H])C(*)=O 0.000 description 1
- 125000000773 L-serino group Chemical group [H]OC(=O)[C@@]([H])(N([H])*)C([H])([H])O[H] 0.000 description 1
- 101100273375 Lactiplantibacillus plantarum (strain ATCC BAA-793 / NCIMB 8826 / WCFS1) cbh gene Proteins 0.000 description 1
- 101100509110 Leifsonia xyli subsp. xyli (strain CTCB07) ispDF gene Proteins 0.000 description 1
- UCOCBWDBHCUPQP-DCAQKATOSA-N Leu-Arg-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(O)=O UCOCBWDBHCUPQP-DCAQKATOSA-N 0.000 description 1
- QDSKNVXKLPQNOJ-GVXVVHGQSA-N Leu-Gln-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O QDSKNVXKLPQNOJ-GVXVVHGQSA-N 0.000 description 1
- OMHLATXVNQSALM-FQUUOJAGSA-N Leu-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(C)C)N OMHLATXVNQSALM-FQUUOJAGSA-N 0.000 description 1
- WMIOEVKKYIMVKI-DCAQKATOSA-N Leu-Pro-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O WMIOEVKKYIMVKI-DCAQKATOSA-N 0.000 description 1
- 241000186780 Listeria ivanovii Species 0.000 description 1
- 241000440393 Listeria monocytogenes EGD-e Species 0.000 description 1
- 108700016271 Listeria monocytogenes actA Proteins 0.000 description 1
- 108700027766 Listeria monocytogenes hlyA Proteins 0.000 description 1
- 101100354184 Listeria monocytogenes serotype 4b (strain F2365) ptcA gene Proteins 0.000 description 1
- 101100426447 Listeria monocytogenes serotype 4b (strain F2365) trpC gene Proteins 0.000 description 1
- 101100098690 Listeria monocytogenes serovar 1/2a (strain ATCC BAA-679 / EGD-e) hly gene Proteins 0.000 description 1
- 241000186807 Listeria seeligeri Species 0.000 description 1
- PNPYKQFJGRFYJE-GUBZILKMSA-N Lys-Ala-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O PNPYKQFJGRFYJE-GUBZILKMSA-N 0.000 description 1
- UWKNTTJNVSYXPC-CIUDSAMLSA-N Lys-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN UWKNTTJNVSYXPC-CIUDSAMLSA-N 0.000 description 1
- CLBGMWIYPYAZPR-AVGNSLFASA-N Lys-Arg-Arg Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O CLBGMWIYPYAZPR-AVGNSLFASA-N 0.000 description 1
- QYOXSYXPHUHOJR-GUBZILKMSA-N Lys-Asn-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O QYOXSYXPHUHOJR-GUBZILKMSA-N 0.000 description 1
- QUYCUALODHJQLK-CIUDSAMLSA-N Lys-Asp-Asp Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O QUYCUALODHJQLK-CIUDSAMLSA-N 0.000 description 1
- IBQMEXQYZMVIFU-SRVKXCTJSA-N Lys-Asp-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCCCN)N IBQMEXQYZMVIFU-SRVKXCTJSA-N 0.000 description 1
- VEGLGAOVLFODGC-GUBZILKMSA-N Lys-Glu-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O VEGLGAOVLFODGC-GUBZILKMSA-N 0.000 description 1
- RFQATBGBLDAKGI-VHSXEESVSA-N Lys-Gly-Pro Chemical compound C1C[C@@H](N(C1)C(=O)CNC(=O)[C@H](CCCCN)N)C(=O)O RFQATBGBLDAKGI-VHSXEESVSA-N 0.000 description 1
- WVJNGSFKBKOKRV-AJNGGQMLSA-N Lys-Leu-Ile Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WVJNGSFKBKOKRV-AJNGGQMLSA-N 0.000 description 1
- YXPJCVNIDDKGOE-MELADBBJSA-N Lys-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CCCCN)N)C(=O)O YXPJCVNIDDKGOE-MELADBBJSA-N 0.000 description 1
- SVSQSPICRKBMSZ-SRVKXCTJSA-N Lys-Pro-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O SVSQSPICRKBMSZ-SRVKXCTJSA-N 0.000 description 1
- HKXSZKJMDBHOTG-CIUDSAMLSA-N Lys-Ser-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CCCCN HKXSZKJMDBHOTG-CIUDSAMLSA-N 0.000 description 1
- QVTDVTONTRSQMF-WDCWCFNPSA-N Lys-Thr-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H]([C@H](O)C)NC(=O)[C@@H](N)CCCCN QVTDVTONTRSQMF-WDCWCFNPSA-N 0.000 description 1
- ZFNYWKHYUMEZDZ-WDSOQIARSA-N Lys-Trp-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CCCCN)N ZFNYWKHYUMEZDZ-WDSOQIARSA-N 0.000 description 1
- 102000011720 Lysophospholipase Human genes 0.000 description 1
- 108020002496 Lysophospholipase Proteins 0.000 description 1
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 1
- 108091006978 Magnesium transporters Proteins 0.000 description 1
- 229920002774 Maltodextrin Polymers 0.000 description 1
- 239000005913 Maltodextrin Substances 0.000 description 1
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 1
- PWHULOQIROXLJO-UHFFFAOYSA-N Manganese Chemical compound [Mn] PWHULOQIROXLJO-UHFFFAOYSA-N 0.000 description 1
- 108010090665 Mannosyl-Glycoprotein Endo-beta-N-Acetylglucosaminidase Proteins 0.000 description 1
- MYAPQOBHGWJZOM-UWVGGRQHSA-N Met-Gly-Leu Chemical compound CSCC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC(C)C MYAPQOBHGWJZOM-UWVGGRQHSA-N 0.000 description 1
- NHXXGBXJTLRGJI-GUBZILKMSA-N Met-Pro-Ser Chemical compound [H]N[C@@H](CCSC)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O NHXXGBXJTLRGJI-GUBZILKMSA-N 0.000 description 1
- 101100124185 Methanococcus maripaludis (strain S2 / LL) hisH1 gene Proteins 0.000 description 1
- 101100327514 Methanosarcina acetivorans (strain ATCC 35395 / DSM 2834 / JCM 12185 / C2A) cfbE gene Proteins 0.000 description 1
- 101100261636 Methanothermobacter marburgensis (strain ATCC BAA-927 / DSM 2133 / JCM 14651 / NBRC 100331 / OCM 82 / Marburg) trpB2 gene Proteins 0.000 description 1
- ZOKXTWBITQBERF-UHFFFAOYSA-N Molybdenum Chemical compound [Mo] ZOKXTWBITQBERF-UHFFFAOYSA-N 0.000 description 1
- 208000035489 Monocytic Acute Leukemia Diseases 0.000 description 1
- 101100063392 Mycobacterium leprae (strain TN) folP1 gene Proteins 0.000 description 1
- 101100293872 Mycolicibacterium smegmatis (strain ATCC 700084 / mc(2)155) sir2 gene Proteins 0.000 description 1
- 102100030397 N-acetylmuramoyl-L-alanine amidase Human genes 0.000 description 1
- 102100023906 N-acylneuraminate-9-phosphatase Human genes 0.000 description 1
- 238000005481 NMR spectroscopy Methods 0.000 description 1
- 108020004485 Nonsense Codon Proteins 0.000 description 1
- 101100276922 Nostoc sp. (strain PCC 7120 / SAG 25.82 / UTEX 2576) dapF2 gene Proteins 0.000 description 1
- 101710138316 O-acetylserine sulfhydrylase Proteins 0.000 description 1
- 206010031096 Oropharyngeal cancer Diseases 0.000 description 1
- 206010057444 Oropharyngeal neoplasm Diseases 0.000 description 1
- 241000178961 Paenibacillus alvei Species 0.000 description 1
- 101100114255 Parasynechococcus marenigrum (strain WH8102) cobQ gene Proteins 0.000 description 1
- 101100338762 Pedobacter heparinus (strain ATCC 13125 / DSM 2366 / CIP 104194 / JCM 7457 / NBRC 12017 / NCIMB 9290 / NRRL B-14731 / HIM 762-3) hepB gene Proteins 0.000 description 1
- 208000002471 Penile Neoplasms Diseases 0.000 description 1
- 108090000279 Peptidyltransferases Proteins 0.000 description 1
- 108010056995 Perforin Proteins 0.000 description 1
- 102000004503 Perforin Human genes 0.000 description 1
- PSKRILMFHNIUAO-JYJNAYRXSA-N Phe-Glu-Lys Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N PSKRILMFHNIUAO-JYJNAYRXSA-N 0.000 description 1
- CWFGECHCRMGPPT-MXAVVETBSA-N Phe-Ile-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O CWFGECHCRMGPPT-MXAVVETBSA-N 0.000 description 1
- WKLMCMXFMQEKCX-SLFFLAALSA-N Phe-Phe-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=CC=C2)NC(=O)[C@H](CC3=CC=CC=C3)N)C(=O)O WKLMCMXFMQEKCX-SLFFLAALSA-N 0.000 description 1
- 102000016462 Phosphate Transport Proteins Human genes 0.000 description 1
- 108010092528 Phosphate Transport Proteins Proteins 0.000 description 1
- 102000015439 Phospholipases Human genes 0.000 description 1
- 108010064785 Phospholipases Proteins 0.000 description 1
- 102100027330 Phosphoribosylaminoimidazole carboxylase Human genes 0.000 description 1
- 108090000434 Phosphoribosylaminoimidazolesuccinocarboxamide synthases Proteins 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- 101100124346 Photorhabdus laumondii subsp. laumondii (strain DSM 15139 / CIP 105565 / TT01) hisCD gene Proteins 0.000 description 1
- 101001135788 Pinus taeda (+)-alpha-pinene synthase, chloroplastic Proteins 0.000 description 1
- 101710183389 Pneumolysin Proteins 0.000 description 1
- CGBYDGAJHSOGFQ-LPEHRKFASA-N Pro-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@@H]2CCCN2 CGBYDGAJHSOGFQ-LPEHRKFASA-N 0.000 description 1
- XZONQWUEBAFQPO-HJGDQZAQSA-N Pro-Gln-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XZONQWUEBAFQPO-HJGDQZAQSA-N 0.000 description 1
- UEHYFUCOGHWASA-HJGDQZAQSA-N Pro-Glu-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H]1CCCN1 UEHYFUCOGHWASA-HJGDQZAQSA-N 0.000 description 1
- RUDOLGWDSKQQFF-DCAQKATOSA-N Pro-Leu-Asn Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O RUDOLGWDSKQQFF-DCAQKATOSA-N 0.000 description 1
- RPLMFKUKFZOTER-AVGNSLFASA-N Pro-Met-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@@H]1CCCN1 RPLMFKUKFZOTER-AVGNSLFASA-N 0.000 description 1
- MHBSUKYVBZVQRW-HJWJTTGWSA-N Pro-Phe-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O MHBSUKYVBZVQRW-HJWJTTGWSA-N 0.000 description 1
- SEZGGSHLMROBFX-CIUDSAMLSA-N Pro-Ser-Gln Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(O)=O SEZGGSHLMROBFX-CIUDSAMLSA-N 0.000 description 1
- OQSGBXGNAFQGGS-CYDGBPFRSA-N Pro-Val-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O OQSGBXGNAFQGGS-CYDGBPFRSA-N 0.000 description 1
- ZMLRZBWCXPQADC-TUAOUCFPSA-N Pro-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@@H]2CCCN2 ZMLRZBWCXPQADC-TUAOUCFPSA-N 0.000 description 1
- 102100034014 Prolyl 3-hydroxylase 3 Human genes 0.000 description 1
- 101710194807 Protective antigen Proteins 0.000 description 1
- 101100405024 Proteus mirabilis (strain HI4320) npdA gene Proteins 0.000 description 1
- 108050008901 Proton-dependent oligopeptide transporter Proteins 0.000 description 1
- 102000000348 Proton-dependent oligopeptide transporter Human genes 0.000 description 1
- 101100070871 Pseudomonas aeruginosa (strain ATCC 15692 / DSM 22644 / CIP 104116 / JCM 14847 / LMG 12228 / 1C / PRS 101 / PAO1) hisH2 gene Proteins 0.000 description 1
- 101100408135 Pseudomonas aeruginosa (strain ATCC 15692 / DSM 22644 / CIP 104116 / JCM 14847 / LMG 12228 / 1C / PRS 101 / PAO1) phnA gene Proteins 0.000 description 1
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 1
- 101100169519 Pyrococcus abyssi (strain GE5 / Orsay) dapAL gene Proteins 0.000 description 1
- 102000009572 RNA Polymerase II Human genes 0.000 description 1
- 108010009460 RNA Polymerase II Proteins 0.000 description 1
- 108020004518 RNA Probes Proteins 0.000 description 1
- 239000003391 RNA probe Substances 0.000 description 1
- 230000006819 RNA synthesis Effects 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 101100313751 Rickettsia conorii (strain ATCC VR-613 / Malish 7) thyX gene Proteins 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 101150006301 SECA2 gene Proteins 0.000 description 1
- MWMKFWJYRRGXOR-ZLUOBGJFSA-N Ser-Ala-Asn Chemical compound N[C@H](C(=O)N[C@H](C(=O)N[C@H](C(=O)O)CC(N)=O)C)CO MWMKFWJYRRGXOR-ZLUOBGJFSA-N 0.000 description 1
- LVVBAKCGXXUHFO-ZLUOBGJFSA-N Ser-Ala-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(O)=O LVVBAKCGXXUHFO-ZLUOBGJFSA-N 0.000 description 1
- WTWGOQRNRFHFQD-JBDRJPRFSA-N Ser-Ala-Ile Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WTWGOQRNRFHFQD-JBDRJPRFSA-N 0.000 description 1
- BRKHVZNDAOMAHX-BIIVOSGPSA-N Ser-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N BRKHVZNDAOMAHX-BIIVOSGPSA-N 0.000 description 1
- BGOWRLSWJCVYAQ-CIUDSAMLSA-N Ser-Asp-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O BGOWRLSWJCVYAQ-CIUDSAMLSA-N 0.000 description 1
- LAFKUZYWNCHOHT-WHFBIAKZSA-N Ser-Glu Chemical compound OC[C@H](N)C(=O)N[C@H](C(O)=O)CCC(O)=O LAFKUZYWNCHOHT-WHFBIAKZSA-N 0.000 description 1
- OHKFXGKHSJKKAL-NRPADANISA-N Ser-Glu-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O OHKFXGKHSJKKAL-NRPADANISA-N 0.000 description 1
- XNCUYZKGQOCOQH-YUMQZZPRSA-N Ser-Leu-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O XNCUYZKGQOCOQH-YUMQZZPRSA-N 0.000 description 1
- ZGFRMNZZTOVBOU-CIUDSAMLSA-N Ser-Met-Gln Chemical compound N[C@@H](CO)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)O ZGFRMNZZTOVBOU-CIUDSAMLSA-N 0.000 description 1
- ADJDNJCSPNFFPI-FXQIFTODSA-N Ser-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CO ADJDNJCSPNFFPI-FXQIFTODSA-N 0.000 description 1
- CKDXFSPMIDSMGV-GUBZILKMSA-N Ser-Pro-Val Chemical compound [H]N[C@@H](CO)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=O CKDXFSPMIDSMGV-GUBZILKMSA-N 0.000 description 1
- HSWXBJCBYSWBPT-GUBZILKMSA-N Ser-Val-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CO)C(C)C)C(O)=O HSWXBJCBYSWBPT-GUBZILKMSA-N 0.000 description 1
- 240000003801 Sigesbeckia orientalis Species 0.000 description 1
- 235000003407 Sigesbeckia orientalis Nutrition 0.000 description 1
- 108091081024 Start codon Proteins 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- 241000194017 Streptococcus Species 0.000 description 1
- 101100278777 Streptomyces coelicolor (strain ATCC BAA-471 / A3(2) / M145) dxs1 gene Proteins 0.000 description 1
- 101100125907 Streptomyces coelicolor (strain ATCC BAA-471 / A3(2) / M145) ilvC1 gene Proteins 0.000 description 1
- 101100126492 Streptomyces coelicolor (strain ATCC BAA-471 / A3(2) / M145) ispG1 gene Proteins 0.000 description 1
- 241000187392 Streptomyces griseus Species 0.000 description 1
- 101100116197 Streptomyces lavendulae dcsC gene Proteins 0.000 description 1
- 101100116199 Streptomyces lavendulae dcsE gene Proteins 0.000 description 1
- 108050004141 Sugar phosphate transporters Proteins 0.000 description 1
- 102000015898 Sugar phosphate transporters Human genes 0.000 description 1
- 108010091582 Sulfate Transporters Proteins 0.000 description 1
- 102000018509 Sulfate Transporters Human genes 0.000 description 1
- 230000024932 T cell mediated immunity Effects 0.000 description 1
- 102220538247 TATA box-binding protein-associated factor RNA polymerase I subunit C_S44A_mutation Human genes 0.000 description 1
- 101100057034 Talaromyces wortmannii astB gene Proteins 0.000 description 1
- JZRWCGZRTZMZEH-UHFFFAOYSA-N Thiamine Natural products CC1=C(CCO)SC=[N+]1CC1=CN=C(C)N=C1N JZRWCGZRTZMZEH-UHFFFAOYSA-N 0.000 description 1
- DFTCYYILCSQGIZ-GCJQMDKQSA-N Thr-Ala-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(O)=O DFTCYYILCSQGIZ-GCJQMDKQSA-N 0.000 description 1
- DWYAUVCQDTZIJI-VZFHVOOUSA-N Thr-Ala-Ser Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O DWYAUVCQDTZIJI-VZFHVOOUSA-N 0.000 description 1
- MPUMPERGHHJGRP-WEDXCCLWSA-N Thr-Gly-Lys Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)N[C@@H](CCCCN)C(=O)O)N)O MPUMPERGHHJGRP-WEDXCCLWSA-N 0.000 description 1
- WFAUDCSNCWJJAA-KXNHARMFSA-N Thr-Lys-Pro Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N1CCC[C@@H]1C(O)=O WFAUDCSNCWJJAA-KXNHARMFSA-N 0.000 description 1
- VTMGKRABARCZAX-OSUNSFLBSA-N Thr-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)[C@@H](C)O VTMGKRABARCZAX-OSUNSFLBSA-N 0.000 description 1
- OLFOOYQTTQSSRK-UNQGMJICSA-N Thr-Pro-Phe Chemical compound C[C@@H](O)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 OLFOOYQTTQSSRK-UNQGMJICSA-N 0.000 description 1
- KZTLZZQTJMCGIP-ZJDVBMNYSA-N Thr-Val-Thr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O KZTLZZQTJMCGIP-ZJDVBMNYSA-N 0.000 description 1
- 108700009124 Transcription Initiation Site Proteins 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 102000002501 Tryptophan-tRNA Ligase Human genes 0.000 description 1
- 102000001742 Tumor Suppressor Proteins Human genes 0.000 description 1
- 108010040002 Tumor Suppressor Proteins Proteins 0.000 description 1
- 101710187743 Tumor necrosis factor receptor superfamily member 1A Proteins 0.000 description 1
- LFTYTUAZOPRMMI-CFRASDGPSA-N UDP-N-acetyl-alpha-D-glucosamine Chemical compound O1[C@H](CO)[C@@H](O)[C@H](O)[C@@H](NC(=O)C)[C@H]1OP(O)(=O)OP(O)(=O)OC[C@@H]1[C@@H](O)[C@@H](O)[C@H](N2C(NC(=O)C=C2)=O)O1 LFTYTUAZOPRMMI-CFRASDGPSA-N 0.000 description 1
- LFTYTUAZOPRMMI-UHFFFAOYSA-N UNPD164450 Natural products O1C(CO)C(O)C(O)C(NC(=O)C)C1OP(O)(=O)OP(O)(=O)OCC1C(O)C(O)C(N2C(NC(=O)C=C2)=O)O1 LFTYTUAZOPRMMI-UHFFFAOYSA-N 0.000 description 1
- 108020000963 Uroporphyrinogen-III synthase Proteins 0.000 description 1
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 description 1
- 241000700618 Vaccinia virus Species 0.000 description 1
- RUCNAYOMFXRIKJ-DCAQKATOSA-N Val-Ala-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN RUCNAYOMFXRIKJ-DCAQKATOSA-N 0.000 description 1
- SYOMXKPPFZRELL-ONGXEEELSA-N Val-Gly-Lys Chemical compound CC(C)[C@@H](C(=O)NCC(=O)N[C@@H](CCCCN)C(=O)O)N SYOMXKPPFZRELL-ONGXEEELSA-N 0.000 description 1
- DLRZGNXCXUGIDG-KKHAAJSZSA-N Val-Thr-Asp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](C(C)C)N)O DLRZGNXCXUGIDG-KKHAAJSZSA-N 0.000 description 1
- WUFHZIRMAZZWRS-OSUNSFLBSA-N Val-Thr-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](C(C)C)N WUFHZIRMAZZWRS-OSUNSFLBSA-N 0.000 description 1
- GVNLOVJNNDZUHS-RHYQMDGZSA-N Val-Thr-Lys Chemical compound [H]N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(O)=O GVNLOVJNNDZUHS-RHYQMDGZSA-N 0.000 description 1
- ZLNYBMWGPOKSLW-LSJOCFKGSA-N Val-Val-Asp Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O ZLNYBMWGPOKSLW-LSJOCFKGSA-N 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 208000004354 Vulvar Neoplasms Diseases 0.000 description 1
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 description 1
- 108091006550 Zinc transporters Proteins 0.000 description 1
- JBGSPJNXVJCLOQ-HJZCUYRDSA-N [(2-amino-2-oxoethyl)-[(3R,4S,5R)-3,4-dihydroxy-5-(hydroxymethyl)oxolane-2-carbonyl]amino]phosphonic acid Chemical compound NC(=O)CN(P(O)(O)=O)C(=O)C1O[C@H](CO)[C@@H](O)[C@H]1O JBGSPJNXVJCLOQ-HJZCUYRDSA-N 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 230000002159 abnormal effect Effects 0.000 description 1
- 230000005856 abnormality Effects 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 230000006978 adaptation Effects 0.000 description 1
- 239000002671 adjuvant Substances 0.000 description 1
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 1
- 108010047495 alanylglycine Proteins 0.000 description 1
- 230000004075 alteration Effects 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 230000005809 anti-tumor immunity Effects 0.000 description 1
- 230000005975 antitumor immune response Effects 0.000 description 1
- 230000006907 apoptotic process Effects 0.000 description 1
- 101150118463 argG gene Proteins 0.000 description 1
- 101150089004 argR gene Proteins 0.000 description 1
- 210000004436 artificial bacterial chromosome Anatomy 0.000 description 1
- 101150057409 asnB gene Proteins 0.000 description 1
- 101150070145 aspB gene Proteins 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 108010047857 aspartylglycine Proteins 0.000 description 1
- 108010068265 aspartyltyrosine Proteins 0.000 description 1
- 101150099905 atpA2 gene Proteins 0.000 description 1
- 101150090348 atpC gene Proteins 0.000 description 1
- 101150099875 atpE gene Proteins 0.000 description 1
- 101150103189 atpG gene Proteins 0.000 description 1
- 229940065181 bacillus anthracis Drugs 0.000 description 1
- 229940097012 bacillus thuringiensis Drugs 0.000 description 1
- UUQMNUMQCIQDMZ-UHFFFAOYSA-N betahistine Chemical compound CNCCC1=CC=CC=N1 UUQMNUMQCIQDMZ-UHFFFAOYSA-N 0.000 description 1
- 229960003237 betaine Drugs 0.000 description 1
- 230000001588 bifunctional effect Effects 0.000 description 1
- 230000008436 biogenesis Effects 0.000 description 1
- 239000012496 blank sample Substances 0.000 description 1
- 101150057791 bsh gene Proteins 0.000 description 1
- 244000309464 bull Species 0.000 description 1
- BNMJSBUIDQYHIN-UHFFFAOYSA-N butyl dihydrogen phosphate Chemical compound CCCCOP(O)(O)=O BNMJSBUIDQYHIN-UHFFFAOYSA-N 0.000 description 1
- 210000004900 c-terminal fragment Anatomy 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 239000011575 calcium Substances 0.000 description 1
- 239000001110 calcium chloride Substances 0.000 description 1
- 229910001628 calcium chloride Inorganic materials 0.000 description 1
- 101150014229 carA gene Proteins 0.000 description 1
- 101150070764 carB gene Proteins 0.000 description 1
- 239000004202 carbamide Substances 0.000 description 1
- 150000001720 carbohydrates Chemical class 0.000 description 1
- 235000014633 carbohydrates Nutrition 0.000 description 1
- 231100000504 carcinogenesis Toxicity 0.000 description 1
- 101150089563 cbiA gene Proteins 0.000 description 1
- 101150038972 cbiD gene Proteins 0.000 description 1
- 230000022131 cell cycle Effects 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 210000000170 cell membrane Anatomy 0.000 description 1
- 201000010881 cervical cancer Diseases 0.000 description 1
- 239000013611 chromosomal DNA Substances 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 239000011436 cob Substances 0.000 description 1
- 101150018117 cobB gene Proteins 0.000 description 1
- 101150103647 cobQ gene Proteins 0.000 description 1
- 101150094543 cobS gene Proteins 0.000 description 1
- 229910017052 cobalt Inorganic materials 0.000 description 1
- 239000010941 cobalt Substances 0.000 description 1
- GUTLYIVDDKVIGB-UHFFFAOYSA-N cobalt atom Chemical compound [Co] GUTLYIVDDKVIGB-UHFFFAOYSA-N 0.000 description 1
- 239000002299 complementary DNA Substances 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 235000009508 confectionery Nutrition 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 238000005138 cryopreservation Methods 0.000 description 1
- 239000013078 crystal Substances 0.000 description 1
- 238000007821 culture assay Methods 0.000 description 1
- 101150094831 cysK gene Proteins 0.000 description 1
- 101150112941 cysK1 gene Proteins 0.000 description 1
- 101150029709 cysM gene Proteins 0.000 description 1
- 230000003436 cytoskeletal effect Effects 0.000 description 1
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 101150011371 dapA gene Proteins 0.000 description 1
- 101150062988 dapF gene Proteins 0.000 description 1
- 101150100742 dapL gene Proteins 0.000 description 1
- 101150012893 dat gene Proteins 0.000 description 1
- 238000007405 data analysis Methods 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 210000004443 dendritic cell Anatomy 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 239000000645 desinfectant Substances 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- ZPTBLXKRQACLCR-XVFCMESISA-N dihydrouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)CC1 ZPTBLXKRQACLCR-XVFCMESISA-N 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- 208000035475 disorder Diseases 0.000 description 1
- 230000003828 downregulation Effects 0.000 description 1
- 101150056470 dxs gene Proteins 0.000 description 1
- 235000013601 eggs Nutrition 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 210000002257 embryonic structure Anatomy 0.000 description 1
- 230000009483 enzymatic pathway Effects 0.000 description 1
- 230000017188 evasion or tolerance of host immune response Effects 0.000 description 1
- 239000013604 expression vector Substances 0.000 description 1
- 101150026389 fabF gene Proteins 0.000 description 1
- 101150035981 fabH gene Proteins 0.000 description 1
- 230000004129 fatty acid metabolism Effects 0.000 description 1
- 101150054723 fliI gene Proteins 0.000 description 1
- 101150073342 folC gene Proteins 0.000 description 1
- 101150045875 folP gene Proteins 0.000 description 1
- 108091008053 gene clusters Proteins 0.000 description 1
- 230000030279 gene silencing Effects 0.000 description 1
- 230000004077 genetic alteration Effects 0.000 description 1
- 231100000118 genetic alteration Toxicity 0.000 description 1
- 238000011331 genomic analysis Methods 0.000 description 1
- 101150041871 gltB gene Proteins 0.000 description 1
- 101150039906 gltD gene Proteins 0.000 description 1
- KWIUHFFTVRNATP-UHFFFAOYSA-N glycine betaine Chemical compound C[N+](C)(C)CC([O-])=O KWIUHFFTVRNATP-UHFFFAOYSA-N 0.000 description 1
- 102000035122 glycosylated proteins Human genes 0.000 description 1
- 108091005608 glycosylated proteins Proteins 0.000 description 1
- VPZXBVLAVMBEQI-UHFFFAOYSA-N glycyl-DL-alpha-alanine Natural products OC(=O)C(C)NC(=O)CN VPZXBVLAVMBEQI-UHFFFAOYSA-N 0.000 description 1
- 108010001064 glycyl-glycyl-glycyl-glycine Proteins 0.000 description 1
- 108010015792 glycyllysine Proteins 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 230000007773 growth pattern Effects 0.000 description 1
- 210000002443 helper t lymphocyte Anatomy 0.000 description 1
- 239000003228 hemolysin Substances 0.000 description 1
- 101150107671 hisB gene Proteins 0.000 description 1
- 101150056694 hisC gene Proteins 0.000 description 1
- 101150113423 hisD gene Proteins 0.000 description 1
- 101150096813 hisF gene Proteins 0.000 description 1
- 101150032598 hisG gene Proteins 0.000 description 1
- 101150091195 hisH gene Proteins 0.000 description 1
- 108010018006 histidylserine Proteins 0.000 description 1
- 101150021605 hlyA gene Proteins 0.000 description 1
- 210000005260 human cell Anatomy 0.000 description 1
- 229910052739 hydrogen Inorganic materials 0.000 description 1
- 101150033780 ilvB gene Proteins 0.000 description 1
- 101150090497 ilvC gene Proteins 0.000 description 1
- 101150043028 ilvD gene Proteins 0.000 description 1
- 101150105723 ilvD1 gene Proteins 0.000 description 1
- 101150077793 ilvH gene Proteins 0.000 description 1
- 101150060643 ilvN gene Proteins 0.000 description 1
- 210000002865 immune cell Anatomy 0.000 description 1
- 230000008629 immune suppression Effects 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 230000002458 infectious effect Effects 0.000 description 1
- 230000008595 infiltration Effects 0.000 description 1
- 238000001764 infiltration Methods 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 238000002347 injection Methods 0.000 description 1
- 239000007924 injection Substances 0.000 description 1
- 101150108670 inlC gene Proteins 0.000 description 1
- 101150067296 inlJ gene Proteins 0.000 description 1
- 239000002054 inoculum Substances 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 150000002506 iron compounds Chemical class 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- 108020001983 isochorismate synthase Proteins 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 101150018742 ispF gene Proteins 0.000 description 1
- 101150081094 ispG gene Proteins 0.000 description 1
- 230000002045 lasting effect Effects 0.000 description 1
- 231100000636 lethal dose Toxicity 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- 230000021633 leukocyte mediated immunity Effects 0.000 description 1
- 230000000670 limiting effect Effects 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 101150082124 lso gene Proteins 0.000 description 1
- 230000002132 lysosomal effect Effects 0.000 description 1
- 108010009298 lysylglutamic acid Proteins 0.000 description 1
- 230000002101 lytic effect Effects 0.000 description 1
- 229910052749 magnesium Inorganic materials 0.000 description 1
- 239000011777 magnesium Substances 0.000 description 1
- 229940035034 maltodextrin Drugs 0.000 description 1
- 229910052748 manganese Inorganic materials 0.000 description 1
- 239000011572 manganese Substances 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 230000001404 mediated effect Effects 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 101150095079 menB gene Proteins 0.000 description 1
- 108020004999 messenger RNA Proteins 0.000 description 1
- 101150095438 metK gene Proteins 0.000 description 1
- 101150115974 metX gene Proteins 0.000 description 1
- 101150043924 metXA gene Proteins 0.000 description 1
- 229960004452 methionine Drugs 0.000 description 1
- 108010022588 methionyl-lysyl-proline Proteins 0.000 description 1
- 101150083023 mgsA gene Proteins 0.000 description 1
- 210000003632 microfilament Anatomy 0.000 description 1
- 238000000386 microscopy Methods 0.000 description 1
- 108091005601 modified peptides Proteins 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 229910052750 molybdenum Inorganic materials 0.000 description 1
- 239000011733 molybdenum Substances 0.000 description 1
- 101150023205 murA1 gene Proteins 0.000 description 1
- 101150089003 murA2 gene Proteins 0.000 description 1
- 101150093075 murD gene Proteins 0.000 description 1
- 101150037968 murG gene Proteins 0.000 description 1
- 101150047269 murI gene Proteins 0.000 description 1
- 231100000707 mutagenic chemical Toxicity 0.000 description 1
- 230000036438 mutation frequency Effects 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 229910052757 nitrogen Inorganic materials 0.000 description 1
- 231100000252 nontoxic Toxicity 0.000 description 1
- 230000003000 nontoxic effect Effects 0.000 description 1
- 230000005257 nucleotidylation Effects 0.000 description 1
- 102000022744 oligopeptide binding proteins Human genes 0.000 description 1
- 108091013547 oligopeptide binding proteins Proteins 0.000 description 1
- 108091008819 oncoproteins Proteins 0.000 description 1
- 102000027450 oncoproteins Human genes 0.000 description 1
- 230000003287 optical effect Effects 0.000 description 1
- 150000002894 organic compounds Chemical class 0.000 description 1
- 201000006958 oropharynx cancer Diseases 0.000 description 1
- 230000002018 overexpression Effects 0.000 description 1
- KHPXUQMNIQBQEV-UHFFFAOYSA-N oxaloacetic acid Chemical compound OC(=O)CC(=O)C(O)=O KHPXUQMNIQBQEV-UHFFFAOYSA-N 0.000 description 1
- 101150013947 pabB gene Proteins 0.000 description 1
- 230000000242 pagocytic effect Effects 0.000 description 1
- 229940014662 pantothenate Drugs 0.000 description 1
- 235000019161 pantothenic acid Nutrition 0.000 description 1
- 239000011713 pantothenic acid Substances 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 108010082406 peptide permease Proteins 0.000 description 1
- 210000005259 peripheral blood Anatomy 0.000 description 1
- 239000011886 peripheral blood Substances 0.000 description 1
- 101150053431 phaZ gene Proteins 0.000 description 1
- 210000001539 phagocyte Anatomy 0.000 description 1
- 101150028857 phoP gene Proteins 0.000 description 1
- 108010088694 phosphoserine aminotransferase Proteins 0.000 description 1
- 102000030592 phosphoserine aminotransferase Human genes 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- 102000054765 polymorphisms of proteins Human genes 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 108010070643 prolylglutamic acid Proteins 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 101150054232 pyrG gene Proteins 0.000 description 1
- 230000005855 radiation Effects 0.000 description 1
- 230000000306 recurrent effect Effects 0.000 description 1
- 239000012926 reference standard material Substances 0.000 description 1
- 230000003252 repetitive effect Effects 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 238000013215 result calculation Methods 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 210000003705 ribosome Anatomy 0.000 description 1
- 101150095594 rluB gene Proteins 0.000 description 1
- 238000010079 rubber tapping Methods 0.000 description 1
- 101150085476 secA1 gene Proteins 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 101150003830 serC gene Proteins 0.000 description 1
- 230000000405 serological effect Effects 0.000 description 1
- 210000002966 serum Anatomy 0.000 description 1
- 239000002356 single layer Substances 0.000 description 1
- 229940063673 spermidine Drugs 0.000 description 1
- 230000006641 stabilisation Effects 0.000 description 1
- 238000011105 stabilization Methods 0.000 description 1
- 239000008227 sterile water for injection Substances 0.000 description 1
- 230000004936 stimulating effect Effects 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 229910052717 sulfur Inorganic materials 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 230000001502 supplementing effect Effects 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 235000019157 thiamine Nutrition 0.000 description 1
- KYMBYSLLVAOCFI-UHFFFAOYSA-N thiamine Chemical compound CC1=C(CCO)SCN1CC1=CN=C(C)N=C1N KYMBYSLLVAOCFI-UHFFFAOYSA-N 0.000 description 1
- 229960003495 thiamine Drugs 0.000 description 1
- 239000011721 thiamine Substances 0.000 description 1
- 101150072448 thrB gene Proteins 0.000 description 1
- 101150000850 thrC gene Proteins 0.000 description 1
- 101150072314 thyA gene Proteins 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 108091006106 transcriptional activators Proteins 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 108091008023 transcriptional regulators Proteins 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 230000001960 triggered effect Effects 0.000 description 1
- 101150019416 trpA gene Proteins 0.000 description 1
- 101150081616 trpB gene Proteins 0.000 description 1
- 101150111232 trpB-1 gene Proteins 0.000 description 1
- 101150044170 trpE gene Proteins 0.000 description 1
- 101150004782 trpS2 gene Proteins 0.000 description 1
- 101150017621 truA gene Proteins 0.000 description 1
- 101150104105 truB gene Proteins 0.000 description 1
- 125000000430 tryptophan group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C2=C([H])C([H])=C([H])C([H])=C12 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 239000000225 tumor suppressor protein Substances 0.000 description 1
- 238000007492 two-way ANOVA Methods 0.000 description 1
- 230000006663 ubiquitin-proteasome pathway Effects 0.000 description 1
- 101150103517 uppS gene Proteins 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 108010063664 uroporphyrin-III C-methyltransferase Proteins 0.000 description 1
- 102000003643 uroporphyrinogen-III synthase Human genes 0.000 description 1
- 210000003934 vacuole Anatomy 0.000 description 1
- 208000013139 vaginal neoplasm Diseases 0.000 description 1
- 230000003612 virological effect Effects 0.000 description 1
- 235000012711 vitamin K3 Nutrition 0.000 description 1
- 239000011652 vitamin K3 Substances 0.000 description 1
- 150000003722 vitamin derivatives Chemical class 0.000 description 1
- 229940041603 vitamin k 3 Drugs 0.000 description 1
- 108010062110 water dikinase pyruvate Proteins 0.000 description 1
- 239000008215 water for injection Substances 0.000 description 1
- 238000002424 x-ray crystallography Methods 0.000 description 1
- 239000011701 zinc Substances 0.000 description 1
- 229910052725 zinc Inorganic materials 0.000 description 1
- 108010088577 zinc-binding protein Proteins 0.000 description 1
Images














Classifications
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5014—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics for testing toxicity
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/315—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Streptococcus (G), e.g. Enterococci
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/02—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving viable microorganisms
- C12Q1/025—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving viable microorganisms for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/02—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving viable microorganisms
- C12Q1/04—Determining presence or kind of microorganism; Use of selective media for testing antibiotics or bacteriocides; Compositions containing a chemical indicator therefor
- C12Q1/06—Quantitative determination
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/483—Physical analysis of biological material
- G01N33/4833—Physical analysis of biological material of solid biological material, e.g. tissue samples, cell cultures
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5044—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics involving specific cell types
- G01N33/5047—Cells of the immune system
- G01N33/5055—Cells of the immune system involving macrophages
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/20011—Papillomaviridae
- C12N2710/20022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Hematology (AREA)
- Urology & Nephrology (AREA)
- Biochemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Biotechnology (AREA)
- Physics & Mathematics (AREA)
- Toxicology (AREA)
- Medicinal Chemistry (AREA)
- Microbiology (AREA)
- Cell Biology (AREA)
- Analytical Chemistry (AREA)
- Genetics & Genomics (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Food Science & Technology (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Biophysics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- General Engineering & Computer Science (AREA)
- Virology (AREA)
- Gastroenterology & Hepatology (AREA)
- Optics & Photonics (AREA)
- Plant Pathology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Peptides Or Proteins (AREA)
Abstract
提供了用于评估细菌或如单核细胞增生性李斯特菌等李斯特菌菌株的减毒和/或感染性的方法和组合物。
Description
相关申请的交叉引用
本申请要求于2018年3月9日提交的美国申请第62/640,855号的权益,所述美国申请出于所有目的通过引用整体并入本文。
对通过EFS WEB以文本文件形式提交的序列表的引用
写入文件528092_SeqListing_ST25.txt中的序列表为89千字节,创建于2019年2月25日,并且通过引用并入本文。
背景技术
单核细胞增生性李斯特菌(Listeria monocytogenes,Lm)是造成人类李斯特菌病的革兰氏阳性、无芽孢形成的细菌生物体。为了使用基于Lm的免疫疗法(如癌症免疫疗法),对细菌进行了生物工程改造以使其减毒,从而可以将其用于递送肿瘤特异性抗原并且产生抗原特异性免疫应答,但不会引起李斯特菌病。原代巨噬细胞可以用于评估基于Lm的免疫疗法感染并在细胞溶质中复制的能力。然而,需要更好的方法来评估李斯特菌菌株的减毒和感染性。
发明内容
提供了用于评估细菌或如单核细胞增生性李斯特菌等李斯特菌菌株的减毒和/或感染性的方法和组合物。在一些方面,提供了用于评估测试李斯特菌菌株的减毒或感染性的方法。此类方法可以包括,例如:(a)用所述测试李斯特菌菌株感染分化的THP-1细胞,其中在用所述测试李斯特菌菌株感染之前,已将所述THP-1细胞分化为巨噬细胞;(b)裂解所述THP-1细胞,并将所述裂解物平板接种在琼脂上;以及;以及(c)通过在所述琼脂上的生长来计数已在所述THP-1细胞内部繁殖的所述李斯特菌。
附图说明
图1A.图展示了资格测定3的绘制为时间对活细胞计数(VCC)的参考标准品和野生型对照的细菌生长速率和倍增时间。
图1B.图展示了资格测定4的绘制为时间对活细胞计数(VCC)的参考标准品和野生型对照的细菌生长速率和倍增时间。
图1C.图展示了资格测定5的绘制为时间对活细胞计数(VCC)的参考标准品和野生型对照的细菌生长速率和倍增时间。
图2A.图展示了绘制为时间对活细胞计数(VCC)的野生型的细菌生长速率和倍增时间,其示出了测定间比较。
图2B.图展示了绘制为时间对活细胞计数(VCC)的参考标准品ADXS11-001的细菌生长速率和倍增时间,其示出了测定间比较。
图3.图展示了在以下时间点观察到的原始计数信息:p-2、p0、p1、p3和p5。
图4.图展示了p-2处的计数与在p0处看到的计数的比率。
图5.图展示了p-2处的计数与在p0处看到的计数的比率,表示为与野生型的比率。
图6.图展示了p3和p%处的计数与在p0处看到的计数的比率。
图7.图展示了相对于野生型,p3和p%处的计数与在p0处看到的计数的比率。
图8.图展示了通过数据运行,相对于野生型,p3和p%处的计数与在p0处看到的计数的比率。
图9.图展示了传代次数相对于野生型从p-2到p0的计数成比例下降的影响。
图10.图展示了回归分析用于评估传代次数的影响。
图11.图展示了图3中每条曲线的两个所得变量之间的关系。
定义
本文可互换使用的术语“蛋白质”、“多肽”、和“肽”是指任何长度的聚合形式的氨基酸,包含编码氨基酸和非编码氨基酸以及以化学方式或生物化学方式修饰的氨基酸或以化学方式或生物化学方式衍生的氨基酸。这些术语包含已经修饰的聚合物,如具有经修饰的肽主链的多肽。
蛋白质被视为具有“N端”和“C端”。术语“N端”是指蛋白质或多肽的起始端,其终止于具有游离胺基(-NH2)的氨基酸。术语“C端”是指氨基酸链(蛋白质或多肽)的末端,其终止于游离羧基(-COOH)。
术语“融合蛋白”是指包括通过肽键或其它化学键连接在一起的两个或更多个肽的蛋白质。所述肽可以通过肽或其它化学键直接连接在一起。例如,可以通过重组方式将嵌合分子表达为单链融合蛋白。可替代地,所述肽可以通过“接头(如一个或多个氨基酸)”或所述两个或更多个肽之间的另一种合适的接头连接在一起。
本文可互换使用的术语“核酸”和“多核苷酸”是指任何长度的聚合形式的核苷酸,包含核糖核苷酸、脱氧核糖核苷酸、或其类似物或经过修饰的版本。所述核苷酸包含单链、双链和多链DNA或RNA、基因组DNA、cDNA、DNA-RNA杂交体以及包括嘌呤碱基、嘧啶碱基或其它天然的、以化学方式修饰的、以生物化学方式修饰的、非天然的或衍生的核苷酸碱基的聚合物。
核酸被视为具有“5'端”和“3'端”,因为以使得一个单核苷酸戊糖环的5'磷酸通过磷酸二酯键在一个方向上与其相邻的单核苷酸戊糖环的3'氧相连的方式使单核苷酸反应以形成寡核苷酸。如果寡核苷酸的5'磷酸不与单核苷酸戊糖环的3'氧相连,则将寡核苷酸的一端称为“5'端”。如果寡核苷酸的3'氧不与另一个单核苷酸戊糖环的5'磷酸相连,则将寡核苷酸的一端称为“3'端”。即使核酸序列处于更大的寡核苷酸的内部,所述核酸序列也可以被视为具有5'端和3'端。在线性或环状DNA分子中,离散元件被称为“下游”或3'元件的“上游”或5'。
“密码子优化”是指通过用宿主细胞的基因中更频繁或最频繁使用的密码子替换天然序列的至少一个密码子同时保持天然氨基酸序列来修饰核酸序列以在特定宿主细胞中增强表达的过程。例如,与天然存在的核酸序列相比,可以修饰对融合多肽进行编码的多核苷酸以取代在给定李斯特菌细胞或任何其它宿主细胞中具有更高使用频率的密码子。密码子使用表在例如“密码子使用数据库”处很容易获得。US 2007/0207170示出了由单核细胞增生性李斯特菌对每个氨基酸利用的最佳密码子,所述文献出于所有目的通过引用整体并入本文。这些表可以通过多种方式进行修改。参见Nakamura等人(2000)《核酸研究(Nucleic Acids Research)》28:292,所述文献出于所有目的通过引用整体并入本文。也可获得用于在特定宿主中表达的特定序列的密码子优化的计算机算法(参见,例如《基因伪造(Gene Forge)》)。
术语“质粒”或“载体”包含任何已知的递送载体,包含细菌递送载体、病毒载体递送载体、肽免疫治疗递送载体、DNA免疫治疗递送载体、附加型质粒、整合型质粒或噬菌体载体。术语“载体”是指能够在宿主细胞中递送和(任选地)表达一种或多种融合多肽的构建体。
术语“附加型质粒”或“染色体外质粒”是指与染色体DNA物理分离(即,附加型或染色体外的,并且未整合到宿主细胞的基因组中)并且独立于染色体DNA复制的核酸载体。质粒可以是线性或环状的,并且可以是单链或双链的。附加型质粒可以任选地在宿主细胞的细胞质(例如,李斯特菌)中以多个拷贝的形式持续存在,从而导致附加型质粒内的所关注的任何基因的扩增。
术语“基因组整合的”是指已被引入到细胞中使得核苷酸序列整合到细胞的基因组中并且能够被其后代遗传的核酸。可以将任何方案用于将核酸稳定地掺入到细胞的基因组中。
术语“稳定保持”是指在不存在选择(例如,抗生素选择)的情况下将核酸分子或质粒保持至少10代而没有可检测到的损失。例如,周期可以是至少15代、20代、至少25代、至少30代、至少40代、至少50代、至少60代、至少80代、至少100代、至少150代、至少200代、至少300代或至少500代。稳定保持可以指核酸分子或质粒在体外(例如,在培养物中)在细胞中稳定地保持、在体内稳定地保持或两者。
“开放阅读框”或“ORF”是含有可能对蛋白质进行编码的碱基序列的DNA的一部分。作为实例,ORF可以定位在基因的起始码序列(起始密码子)与终止密码子序列(终止密码子)之间。
“启动子”是DNA的调节区域,其通常包括能够指导RNA聚合酶II在特定多核苷酸序列的适当转录起始位点处起始RNA合成的TATA盒。启动子可以另外包括影响转录起始速率的其它区域。本文所公开的启动子序列调节可操作连接的多核苷酸的转录。启动子可以在本文所公开的细胞类型(例如,真核细胞、非人类哺乳动物细胞、人类细胞、啮齿动物细胞、多能细胞、单细胞阶段胚胎、分化细胞或其组合)中的一种或多种细胞类型中具有活性。启动子可以是例如组成型活性启动子、条件型启动子、诱导型启动子、时间受限启动子(例如,受发育调节的启动子)或空间受限启动子(例如,细胞特异性或组织特异性启动子)。启动子的实例可以例如在WO 2013/176772中找到,所述文献通过引用整体并入本文。
“可操作的连接”或“可操作地连接”是指将两种或多种组分(例如启动子和另一种序列元件)并置使得两种组分正常发挥功能,并使得至少一种组分能够介导施加在至少一种其它组分上的功能。例如,如果启动子响应于存在或不存在一种或多种转录调节因子而控制编码序列的转录水平,则可将所述启动子可操作地连接到编码序列。可操作的连接可以包含这些序列彼此相邻或以反式作用(例如,调节序列可在一定距离处起作用以控制编码序列的转录)。
“序列同一性”或“同一性”在两个多核苷酸或多肽序列的上下文中是指当在指定的比较窗口上出于最大对应进行比对时两个序列中相同的残基。当提及蛋白质的序列同一性的百分比时,认识到不相同的残基位置通常因保守性氨基酸取代而不同,其中氨基酸残基被具有相似化学性质(例如,电荷或疏水性)的其它氨基酸残基取代,因此不改变分子的功能性质。当序列的保守性取代不同时,可以将百分比序列同一性向上调整以校正取代的保守性质。因这些保守性取代而不同的序列被视为具有“序列相似性”或“相似性。”用于进行这种调整的方法众所周知。通常,这涉及将保守性取代计为部分错配而不是完全错配,从而增加百分比序列同一性。因此,例如,当相同氨基酸的所得评分为1,非保守性取代的所得评分为零时,保守性取代的所得评分介于零与1之间。例如,通过在项目PC/GENE(加利福尼亚州山景城的Intelligenetics公司(Intelligenetics,Mountain View,California))中的实施方式计算保守性取代的评分。
“序列同一性百分比”是指通过在比较窗口上比较两个最佳比对序列确定的值(完全匹配残基的最大数量),其中在比较窗口中的多核苷酸序列部分与参考序列(不包括添加物或缺失部分)相比可以包括添加物或缺失部分(即缺口),以实现两个序列的最佳比对。通过确定在两个序列中出现相同核酸碱基或氨基酸残基的位置数计算百分比来得到匹配位置数,用匹配位置数除以比较窗口中的位置总数,并将结果乘以100以得到序列同一性的百分比。除非另有说明(例如较短的序列包含连接的异源序列),否则所述比较窗口为两个所比较序列中较短序列的全长。
除非另有说明,否则序列同一性/相似性值是指使用以下参数使用第10版GAP获得的值:使用GAP权重50、长度权重3以及nwsgapdna.cmp评分矩阵的核苷酸序列的同一性百分比和相似性百分比;使用GAP权重8和长度权重2以及BLOSUM62评分矩阵的氨基酸序列的同一性百分比和相似性百分比;或其任何等效程序。“等效程序”包含当与第10版GAP生成的相应比对进行比较时针对所讨论的任何两个序列产生具有相同核苷酸或氨基酸残基匹配和相同百分比序列同一性的比对的任何序列比较程序。
术语“保守性氨基酸取代”是指用具有相似大小、电荷或极性的不同氨基酸取代序列中正常存在的氨基酸。保守性取代的实例包含用非极性(疏水性)残基(如异亮氨酸、缬氨酸或亮氨酸)取代另一种非极性残基。类似地,保守性取代的实例包含用一种极性(亲水性)残基取代另一种极性残基,如精氨酸与赖氨酸之间的极性残基、谷氨酰胺与天冬酰胺之间的极性残基或甘氨酸与丝氨酸之间的极性残基。此外,用碱性残基(如赖氨酸、精氨酸或组氨酸)取代另一种碱性残基或者用一种酸性残基(如天冬氨酸或谷氨酸)取代另一种酸性残基是保守性取代另外的实例。非保守性取代的实例包含用非极性(疏水性)氨基酸残基(如异亮氨酸、缬氨酸、亮氨酸、丙氨酸或甲硫氨酸)取代极性(亲水性)残基(如半胱氨酸、谷氨酰胺、谷氨酸或赖氨酸)和/或用极性残基取代非极性残基。典型的氨基酸分类总结如下。
表1.氨基酸分类。

“同源”序列(例如,核酸序列)是指与已知参考序列相同或基本相似的序列,使得其与已知参考序列具有例如至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%或100%同一性。
术语“野生型”是指具有在正常(与突变、患病、改变等相比)状态或情况下发现的结构和/或活性的实体。野生型基因和多肽通常以多种不同形式(例如,等位基因)存在。
关于蛋白质和核酸的术语“分离的”是指相对于通常原位存在、达到并且包含蛋白质和多核苷酸的基本上纯的调配物的其它细菌、病毒或细胞组分相对纯化的蛋白质和核酸。术语“分离的”还包含不具有天然存在的对应物、已经被化学合成并且因此基本上未被其它蛋白质或核酸污染或者已经与其天然伴随有的大多数其它细胞组分(例如,其它细胞蛋白、多核苷酸或细胞组分)分离或纯化的蛋白质或核酸。
“外源”或“异源”分子或序列是通常在细胞中不表达或通常不以所述形式存在于细胞中的分子或序列。正常存在包含关于细胞的特定发育阶段和环境条件的存在。例如,外源或异源性分子或序列可以包含细胞内相应的内源序列的突变版本,或者可以包含与细胞内的内源序列相对应但形式不同(即,不在染色体内)的序列。特定细胞中的外源或异源性分子或序列也可以是源自与细胞的参考物种不同的物种或源自同一物种内的不同生物体的分子或序列。例如,在表达异源多肽的李斯特菌菌株的情况下,异源多肽可以是非李斯特菌菌株天然或内源的、不由李斯特菌菌株正常表达的、来自除李斯特菌菌株之外的来源、源自同一物种内的不同生物体的多肽。
相比之下,“内源”分子或序列或“天然”分子或序列是在特定环境条件下在特定发育阶段在特定细胞中通常以所述形式存在的分子或序列。
术语“变体”是指不同于大多数群体,但仍与被认为是其中之一的通用模式足够相似的氨基酸或核酸序列(或生物体或组织)(例如,剪接变体)。
术语“同工型”是指分子(例如,蛋白质)的版本,其与(例如,同一蛋白质的)另一个同工型或版本相比仅具有微小差异。例如,蛋白质同工型可以从不同但有关的基因中产生,其可以通过替代性剪接从同一基因中产生,或者其可以从单核苷酸多态性中产生。
当提及蛋白质时,术语“片段”意指比全长蛋白质更短或具有更少氨基酸的蛋白质。当提及核酸时,术语“片段”意指比全长核酸更短或具有更少核苷酸的核酸。片段可以是例如N端片段(即,去除蛋白质的C端的一部分)、C端片段(即,去除蛋白质的N端的一部分)或内部片段。片段也可以是例如功能性片段或免疫原性片段。
当涉及蛋白质时,术语“类似物”意指通过保守性氨基酸差异、通过不影响氨基酸序列的修饰或通过两者而与天然存在的蛋白质不同的蛋白质。
术语“功能性”是指蛋白质或核酸(或其片段、同工型或变体)表现出生物活性或功能的先天能力。此类生物活性或功能可以包含例如当施用于受试者时引起免疫应答的能力。此类生物活性或功能还可以包含例如与相互作用配偶体的结合。在功能性片段、同工型或变体的情况下,这些生物功能实际上可以发生改变(例如,关于其特异性或选择性),但保留基本生物功能。
术语“免疫原性”或“免疫原性的”是指分子(例如,蛋白质、核酸、抗原或生物体)在施用于受试者时引起受试者免疫应答的先天能力。免疫原性可以例如通过针对分子的更多数量的抗体、针对分子的更多种类的抗体、对分子具有特异性的更多数量的T细胞、针对分子的更大的细胞毒性或辅助T细胞应答等来测量。
在本文中,术语“抗原”用于指当与受试者或生物体接触时(例如,当存在于受试者或生物体中或由所述受试者或生物体检测到时)使得所述受试者或生物体产生可检测的免疫应答的物质。抗原可以是例如脂质、蛋白质、碳水化合物、核酸或其组合和变体。例如,“抗原肽”是指当存在于受试者或生物体中或由所述受试者或生物体检测到时使得所述受试者或生物体中的免疫应答增加的肽。例如,这种“抗原肽”可以涵盖装载并呈现在宿主细胞表面上的MHC I类和/或II类分子上的蛋白质,并且可以由宿主的免疫细胞识别或检测,从而使得针对所述蛋白质的免疫应答增加。这种免疫应答也可以扩展到宿主内的其它细胞,如表达相同蛋白质的患病细胞(例如,肿瘤或癌细胞)。
术语“表位”是指抗原上被免疫系统识别的位点(例如,抗体结合的位点)。表位可以由连续氨基酸或通过一种或多种蛋白质的三级折叠而并置的非连续氨基酸形成。由连续氨基酸形成的表位(也被称为线性表位)通常在暴露于变性溶剂时保留,而由三级折叠形成的表位(也被称为构象表位)通常在用变性溶剂处理时丢失。在独特的空间构象中,表位典型地包含至少3个(并且更常见地,至少5个或8-10个)氨基酸。确定表位的空间构象的方法包含例如X射线晶体学和2维核磁共振。参见例如,Glenn E.Morris编辑的《分子生物学方法(Methods in Molecular Biology)》第66卷中的《表位作图指南(Epitope MappingProtocols)》(1996),所述文献出于所有目的通过引用整体并入本文。
术语“突变”是指基因或蛋白质的结构的任何改变。例如,突变可以由染色体或蛋白质的缺失、插入、取代或重排引起。“插入”通过添加一个或多个另外的核苷酸或氨基酸来改变基因中的核苷酸的数量或蛋白质中的氨基酸的数量。“缺失”通过减少一个或多个另外的核苷酸或氨基酸来改变基因中的核苷酸的数量或蛋白质中的氨基酸的数量。
当核苷酸的添加或缺失改变基因的阅读框时,DNA就会发生“移码”突变。阅读框由3个碱基的组组成,每个碱基编码一个氨基酸。移码突变转变这些碱基的分组,并且改变氨基酸代码。所得的蛋白质通常是无功能的。插入和缺失均可以是移码突变。
“错义”突变或取代是指蛋白质的一个氨基酸的改变或单个核苷酸中的点突变,其导致经编码的氨基酸改变。导致一个氨基酸改变的单个核苷酸中的点突变是DNA序列中的“非同义”取代。非同义取代也可能导致“无意义”突变,其中密码子更改为提前终止密码子,所述提前终止密码子导致所得的蛋白质被截短。相比之下,DNA中的“同义”突变是不更改蛋白质的氨基酸序列的突变(由于密码子简并性)。
术语“体细胞突变”包含由生殖细胞以外的细胞(例如,精子或卵)获得的遗传学改变。此类突变可以在细胞分裂的过程中传递给突变细胞的后代,但不可遗传。相比之下,生殖细胞突变发生在种系中,并且可以传递给下一代后代。
术语“体外(in vitro)”是指人工环境以及在人工环境(例如,试管)内发生的过程或反应。
术语“体内(in vivo)”是指自然环境(例如,细胞、生物体或身体)以及在自然环境内发生的过程或反应。
“包括”或“包含”一个或多个所列举的元件的组合物或方法可以包含其它未具体列举的元件。例如,“包括”或“包含”蛋白质的组合物可以单独含有蛋白质或与其它成分组合的蛋白质。
数值范围的指定包含所述范围内或定义所述范围的所有整数以及由所述范围内的整数定义的所有子范围。
除非从上下文中明显看出,否则术语“约”涵盖规定值的标准测量误差范围(例如,SEM)内的值或与指定值相差±0.5%、±1%、±5%或±10%的值。
除非上下文另外明确指明,否则本文中的单数形式“一个/一种(a/an)”和“所述(the)”包含复数个提及物。例如,术语“抗原”或“至少一种抗原”可以包含多种抗原,包含其混合物。
统计学上显著意指p≤0.05。
具体实施方式
I.概述
本文公开了使用分化的THP-1细胞来分析基于李斯特菌的免疫疗法的细胞内生长的基于细胞的测定法。此类测定可以用于例如评估与野生型李斯特菌相比重组李斯特菌菌株的减毒或评估重组李斯特菌菌株的效力或感染性。
作为一个具体实例,ADXS11-001是由于基因组中的prfA的不可逆缺失以及其与突变的prfA基因(D133V)的互补而减毒的重组单核细胞增生性李斯特菌(Lm)菌株。prfA基因调节体内细胞内生长和Lm存活所需的几种毒力基因(如hly(李斯特菌溶血素O或LLO)、actA(肌动蛋白成核剂A)、plcA(磷脂酶A)和plcB(磷脂酶B))的转录。在ADXS11-001中与突变的prfA的互补导致毒力基因表达的降低。ADXS11-001免疫疗法中的质粒还含有在hly启动子控制下与截短的李斯特菌溶血素O(tLLO)融合的人乳头瘤病毒蛋白E7。为了评估ADXS11-001的减毒,使用野生型Lm作为对照,在巨噬细胞感染测定中评估感染和复制。
ADXS11-001的生物活性取决于抗原呈递细胞(APC)(如巨噬细胞和树突状细胞)对ADXS11-001的摄取、抗原呈递细胞从吞噬溶酶体中逃逸、APC的细胞溶质中的细胞内复制、tLLO-E7的表达、加工以及tLLO-E7在APC的表面上的呈递,以刺激E7特异性细胞毒性T细胞应答。使用分化的THP-1细胞是替代使用原代巨噬细胞监测ADXS11-001在巨噬细胞的细胞溶质中感染和复制的能力的更好的方案。该方法的优点还在于其是定量的。
II.用于评估李斯特菌的减毒和感染性的方法
提供了用于评估细菌的减毒和/或感染性的方法和组合物。在一些实施例中,所述细菌是李斯特菌菌株。在一些实施例中,所述李斯特菌菌株是单核细胞增生性李斯特菌菌株。在一些实施例中,所述单核细胞增生性李斯特菌菌株是突变、重组或减毒的单核细胞增生性李斯特菌菌株。可以用于此类方法的重组李斯特菌菌株的实例在本文其它地方更详细地提供。此类方法利用具有巨噬细胞表型的巨噬细胞细胞系或类巨噬细胞细胞系。此类细胞可以是永生化细胞。例如,细胞系可以是人单核细胞系,如THP-1细胞。THP-1指自发永生化单核样细胞系,其源自儿童急性单核细胞白血病(M5亚型)的外周血。可以使用例如佛波醇12-肉豆蔻酸酯13-乙酸酯(通常被称为PMA或TPA)将THP-1细胞分化为巨噬细胞样细胞。
在一些实施例中,所述方法包括:(a)用测试李斯特菌菌株感染分化的THP-1细胞,其中在用所述测试李斯特菌菌株感染之前,已将所述THP-1细胞分化为巨噬细胞;(b)裂解所述THP-1细胞,并将所述裂解物平板接种在琼脂上;以及(c)通过在所述琼脂上的生长来计数已在所述THP-1细胞内部繁殖的所述李斯特菌。分化的THP-1细胞可以作为粘附细胞生长。也可以使用其它巨噬细胞样细胞。也可以使用其它巨噬细胞样永生化细胞和/或细胞系。
在一些实施例中,所述方法进一步包括将所述THP-1细胞分化为巨噬细胞。例如,如本文其它地方所公开的,可以在步骤(a)之前使用佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA)来完成这种分化。在一些实施例中,在分化之前,THP-1细胞的传代次数小于32。
在一些实施例中,步骤(a)包括以1:1的感染复数(MOI)感染所述分化的THP-1细胞。然而,可以使用任何合适的感染复数。
任选地,此类方法可以进一步包括在步骤(a)与(b)之间杀死未被所述THP-1细胞摄取的李斯特菌。例如,可以使用抗生素(如庆大霉素)进行杀死。
任选地,裂解步骤(b)在感染后3小时进行。然而,裂解步骤也可以在其它时间点(如感染后1、2、3、4、5、6、7、8、9或10小时)进行。
在一些实施例中,用细菌菌株感染分化的THP-1细胞包括将细菌与分化的THP-1细胞一起孵育1-5小时、2-3小时、1小时、2小时、3小时、2小时±60分钟、2小时±50分钟、2小时±40分钟、2小时±30分钟、2小时±25分钟、2小时±20分钟、2小时±15分钟、2小时±10分钟、2小时±5分钟或2小时±3分钟。在一些实施例中,所述细菌是李斯特菌。在一些实施例中,所述李斯特菌是单核细胞增生性李斯特菌。在一些实施例中,相对于野生型单核细胞增生性李斯特菌,所述单核细胞增生性李斯特菌是减毒的。在一些实施例中,将含有细菌的接种培养基添加至分化的THP-1细胞。
在一些实施例中,感染步骤进一步包括一个或多个洗涤步骤和/或杀死步骤。洗涤步骤可以包括从THP-1细胞中去除含细菌的培养基,并且任选地冲洗THP-1细胞,从而去除尚未感染THP-1细胞的细菌。如果使用洗涤步骤,则可以在细菌与THP-1细胞孵育之后和裂解步骤之前进行洗涤步骤。杀死步骤可以包括将对细菌有效的抗生素添加至THP-1细胞,从而杀死未被THP-1细胞摄取的细菌(即,细胞外细菌)。可以以有效杀死细菌的浓度添加抗生素。如果使用杀死步骤,则可以在细菌与THP-1细胞孵育之后和裂解步骤之前进行杀死步骤。杀死步骤可以在洗涤步骤之后或之前进行,或在两个洗涤步骤之间进行。在一些实施例中,将抗生素添加至THP-1细胞并孵育15-75分钟、20-60分钟、30-50分钟或约42-45分钟。在一些实施例中,抗生素是庆大霉素。
在一些实施例中,在感染步骤之后(感染后0小时)、感染后0-10小时、感染后1小时、感染后2小时、感染后3小时、感染后4小时、感染后5小时、感染后6小时、感染后7小时、感染后8小时、感染后9小时或感染后10小时立即进行裂解步骤(b)。在一些实施例中,在感染步骤之后(p0)、感染后1小时(p1)、感染后3小时(p3)或感染后5小时(p5)立即进行裂解步骤。如果在感染步骤之后未立即进行裂解,则可以将THP-1细胞在生长培养基中孵育直至裂解。感染后的孵育期间可能发生细菌的细胞内生长。裂解步骤可以包括将THP-1细胞收集在能够裂解THP-1细胞而不是细菌的水或类似溶剂中以形成裂解物,并且将裂解物平板接种在能够支持细菌生长并允许计数菌落形成单位(CFU)的数量的培养基上。在一些实施例中,可以将裂解物稀释。在一些实施例中,可以将裂解物的一种或多种不同的稀释液平板接种在培养基上。
在一些实施例中,计数步骤可以包括确定来自裂解物的CFU的数量。在一些实施例中,确定接种培养基中的CFU的数量。在一些实施例中,在不同的感染后裂解期或细菌菌株之后确定CFU的数量。在一些实施例中,在感染步骤之后即刻以及感染后一个或多个时间针对接种培养基确定细菌菌株的CFU。在一些实施例中,在感染步骤之后和感染后三小时立即确定细菌菌株的CFU。在一些实施例中,在一个时间确定CFU并且与在另一感染后时间确定的CFU进行比较。在一些实施例中,通过将接种培养基的CFU与感染后0小时的CFU进行比较来计算摄取或感染速率。在一些实施例中,通过将感染后1-10小时的CFU与感染后0小时的CFU进行比较来计算细胞内生长速率。在一些实施例中,通过将感染后1小时、3小时或5小时的CFU与感染后0小时确定的CFU进行比较来计算细胞内生长速率。
此类方法可以进一步包括将测试细菌菌株(如突变、重组或减毒的单核细胞增生性李斯特菌菌株)的摄取和/或细胞内生长与对照(如野生型李斯特菌菌株)和/或参考样品进行比较。
在实例中公开了另外的实施例。
III.重组细菌或李斯特菌菌株
本文公开的方法评估细菌菌株(如李斯特菌菌株)的减毒和感染性。此类细菌菌株可以是重组细菌菌株。此类重组细菌菌株可以包括本文所公开的重组融合多肽或对如本文其它各处所公开的重组融合多肽进行编码的核酸。优选地,细菌菌株是李斯特菌菌株,如单核细胞增生性李斯特菌(Lm)菌株。Lm作为疫苗载体具有许多固有的优势。细菌无需特殊要求即可在体外高效生长,并且缺乏LPS,所述LPS是革兰氏阴性细菌(如沙门氏菌)的主要毒性因子。基因减毒的Lm载体还具有另外的安全性,因为在严重副作用的情况下其可以很容易地用抗生素消除,并且与一些病毒载体不同,遗传物质不会整合到宿主基因组中。
重组李斯特菌菌株可以是任何李斯特菌菌株。合适的李斯特菌菌株的实例包含斯氏李斯特菌(Listeria seeligeri)、格氏李斯特菌(Listeria grayi)、伊氏李斯特菌(Listeria ivanovii)、默氏李斯特菌(Listeria murrayi)、威氏李斯特菌(Listeriawelshimeri)、单核细胞增生性李斯特菌(Lm)或任何其它已知的李斯特菌物种。优选地,重组李斯特菌菌株是单核细胞增生性李斯特菌物种的菌株。单核细胞增生性李斯特菌菌株的实例包含以下:单核细胞增生性李斯特菌10403S野生型(参见例如,Bishop和Hinrichs(1987)《免疫学杂志(J Immunol)》139:2005-2009;Lauer等人(2002)《细菌学杂志(JBact)》184:4177-4186);噬菌体治愈的单核细胞增生性李斯特菌DP-L4056(参见例如,Lauer等人(2002)《细菌学杂志》184:4177-4186);噬菌体治愈的并且具有hly基因缺失的单核细胞增生性李斯特菌DP-L4027(参见例如,Lauer等人(2002《细菌学杂志》184:4177-4186;Jones和Portnoy(1994)《感染与免疫(Infect Immunity)》65:5608-5613;噬菌体治愈的并且具有actA基因缺失的单核细胞增生性李斯特菌DP-L4029(参见例如,Lauer等人(2002《细菌学杂志》184:4177-4186;Skoble等人(2000)《细胞生物学杂志(J Cell Biol)》150:527-538);单核细胞增生性李斯特菌DP-L4042(δPEST)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊(Proc Natl Acad Sci.USA)》101:13832-13837以及支持信息);单核细胞增生性李斯特菌DP-L4097(LLO-S44A)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌DP-L4364(δlplA;硫辛酸酯蛋白连接酶)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌DP-L4405(δinlA)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌DP-L4406(δinlB)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌CS-LOOOl(δactA;δinlB)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌CS-L0002(δactA;δlplA)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌CS-L0003(LLO L461T;δlplA)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌DP-L4038(δactA;LLO L461T)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);单核细胞增生性李斯特菌DP-L4384(LLO S44A;LLO L461T)(参见例如,Brockstedt等人(2004)《美国国家科学院院刊》101:13832-13837以及支持信息);具有lplA1缺失的单核细胞增生性李斯特菌(对硫辛酸酯蛋白连接酶LplA1进行编码)(参见例如,O’Riordan等人(2003)《科学(Science)》302:462-464);单核细胞增生性李斯特菌DP-L4017(具有LLOL461T的10403S)(参见例如,US 7,691,393);单核细胞增生性李斯特菌EGD(参见例如,基因库(GenBank)登录号AL591824)。在另一个实施例中,李斯特菌菌株是单核细胞增生性李斯特菌EGD-e(参见基因库登录号NC_003210;ATCC登录号BAA-679);单核细胞增生性李斯特菌DP-L4029(actA缺失,任选地与uvrAB缺失组合(DP-L4029uvrAB))(参见例如,US 7,691,393);单核细胞增生性李斯特菌actA-/inlB-双突变体(参见例如,ATCC登录号PTA-5562);单核细胞增生性李斯特菌lplA突变体或hly突变体(参见例如,US 2004/0013690);单核细胞增生性李斯特菌dal/dat-双突变体(参见例如,US 2005/0048081)。其它单核细胞增生性李斯特菌菌株包含经修饰(例如,通过质粒和/或通过基因组整合)以含有对以下基因中的一种基因或其任何组合进行编码的核酸的单核细胞增生性李斯特菌菌株:hly(LLO;李斯特菌溶血素);iap(p60);inlA;inlB;inlC;Dal(丙氨酸消旋酶);dat(D-氨基酸氨基转移酶);plcA;plcB;actA;或介导单壁囊泡生长、扩散、分解、双壁囊泡分解、与宿主细胞结合或被宿主细胞摄取的任何核酸。上述参考文献中的每个参考文献出于所有目的通过引用整体并入本文。
重组细菌或李斯特菌可以具有野生型毒力,可以具有减毒的毒力,或者可以是无毒力的。例如,重组的李斯特菌可以具有足够的毒性,以逃避吞噬体或吞噬溶酶体并且进入细胞溶质。这种李斯特菌菌株也可以是活的减毒的李斯特菌菌株,其包括至少一种如本文其它地方所公开的减毒突变、缺失或失活。优选地,重组李斯特菌是减毒的营养缺陷型菌株。营养缺陷型菌株是无法合成其生长所需的特定有机化合物的菌株。此类菌株的实例描述于US 8,114,414中,所述文献出于所有目的通过引用整体并入本文。
优选地,重组李斯特菌菌株缺乏抗生素抗性基因。例如,此类重组李斯特菌菌株可以包括不对抗生素抗性基因进行编码的质粒。然而,本文提供的一些重组李斯特菌菌株包括质粒,所述质粒包括对抗生素抗性基因进行编码的核酸。抗生素抗性基因可以用于分子生物学和疫苗制备中常用的常规选择和克隆过程。示例性抗生素抗性基因包含赋予对氨苄青霉素、青霉素、甲氧西林、链霉素、红霉素、卡那霉素、四环素、氯霉素(CAT)、新霉素、潮霉素和庆大霉素的抗性的基因产物。
A.包括重组融合多肽或对重组融合多肽进行编码的核酸的细菌或李斯特菌菌株
本文所公开的重组细菌菌株(例如,李斯特菌菌株)包括本文所公开的重组融合多肽或对如本文其它各处所公开的重组融合多肽进行编码的核酸。
在包括对重组融合蛋白进行编码的核酸的细菌或李斯特菌菌株中,可以对核酸进行密码子优化。US 2007/0207170示出了由单核细胞增生性李斯特菌对每个氨基酸利用的最佳密码子的实例,所述文献出于所有目的通过引用整体并入本文。如果核酸中的至少一个密码子被替换为由单核细胞增生性李斯特菌针对所述氨基酸比原始序列中的密码子更频繁使用的密码子,则所述核酸是密码子优化的。
核酸可以存在于细菌或李斯特菌菌株内的附加型质粒中和/或核酸可以基因组整合于细菌或李斯特菌菌株中。一些重组细菌或李斯特菌菌株包括对如本文所公开的两个重组融合多肽进行编码的两个单独的核酸:附加型质粒中的一个核酸,以及基因组整合于细菌或李斯特菌菌株中的一个核酸。
附加型质粒可以是在体外(在细胞培养中)、体内(在宿主中)或在体外和体内都稳定保持的质粒。如果在附加型质粒中,则对重组融合多肽进行编码的开放阅读框可以可操作地连接至质粒中的启动子/调节序列。如果基因组整合于细菌或李斯特菌菌株中,则对重组融合多肽进行编码的开放阅读框可以可操作地连接至外源启动子/调节序列或内源启动子/调节序列。用于驱动基因的组成型表达的启动子/调节序列的实例是公知的,并且包含例如李斯特菌的hly、hlyA、actA、prfA和p60启动子、链球菌bac启动子、灰色链霉菌sgiA启动子和苏云金芽孢杆菌phaZ启动子。在一些情况下,插入的所关注的基因不会被中断或受到通常由于整合到基因组DNA中而发生的调节限制,并且在一些情况下,插入的异源基因的存在不会导致细胞自身重要区域的重排或中断。
可以通过用包括对重组融合多肽进行编码的核酸的质粒或载体转化本文其它地方所描述的细菌或李斯特菌菌株或减毒的细菌或李斯特菌菌株,从而制备此类重组细菌或李斯特菌菌株。质粒可以是不整合到宿主染色体中的附加型质粒。可替代地,质粒可以是整合到细菌或李斯特菌菌株的染色体中的整合型质粒。本文所使用的质粒也可以是多拷贝质粒。用于转化细菌的方法是公知的,并且包含基于氯化钙感受态细胞的方法、电穿孔方法、噬菌体介导的转导、化学转化技术和物理转化技术。参见例如,de Boer等人(1989)《细胞(Cell)》56:641-649;Miller等人(1995)《美国生物实验学学会联合会刊(FASEB J.)》9:190-199;Sambrook等人(1989)《分子克隆:实验室手册(Molecular Cloning:A LaboratoryManual)》,冷泉港实验室(Cold Spring Harbor Laboratory),纽约;Ausubel等人(1997)《当代分子生物学实验指南(Current Protocols in Molecular Biology)》,约翰·威利父子出版公司(John Wiley&Sons),纽约;Gerhardt等人编辑,1994,《通用和分子细菌学方法(Methods for General and Molecular Bacteriology)》,美国微生物学会(AmericanSociety for Microbiology),华盛顿特区(Washington,D.C.);以及Miller,1992,《细菌遗传学短期课程(A Short Course in Bacterial Genetics)》,冷泉港实验室出版社(ColdSpring Harbor Laboratory Press),纽约州冷泉港(Cold Spring Harbor,N.Y.),所述文献中的每个文献出于所有目的通过引用整体并入本文。
具有经基因组整合的异源核酸的细菌或李斯特菌菌株可以例如通过使用位点特异性整合载体来制备,由此使用同源重组产生包括整合基因的细菌或李斯特菌。整合载体可以是能够感染细菌或李斯特菌菌株的任何位点特异性整合载体。此类整合载体可以包括例如PSA attPP'位点、对PSA整合酶进行编码的基因、U153 attPP'位点、对U153整合酶进行编码的基因、A118 attPP'位点、对A118整合酶进行编码的基因、或任何其它已知的attPP'位点或任何其它噬菌体整合酶。
包括整合基因的此类细菌或李斯特菌菌株也可以使用用于将异源核酸整合到细菌或李斯特菌染色体中的任何其它已知方法来产生。同源重组的技术是公知的,并且在以下文献中进行了描述:例如,Baloglu等人(2005)《兽医微生物学(Vet Microbiol)》109(1-2):11-17;Jiang等人(2005)《生物化学与生物物理学杂志(Acta Biochim Biophys Sin)》(上海)37(1):19-24以及US 6,855,320,所述文献中的每个文献出于所有目的通过引用整体并入本文。
也可以使用转座子插入实现整合到细菌或李斯特菌染色体中。转座子插入技术是公知的,并且针对例如DP-L967的构建进行了描述:Sun等人(1990)《感染与免疫(Infectionand Immunity)》58:3770-3778,所述文献出于所有目的通过引用整体并入本文。转座子诱变可以实现稳定的基因组插入,但是基因组中已插入异源核酸的位置是未知的。
也可以使用噬菌体整合位点实现整合到细菌或李斯特菌染色体中(参见例如,Lauer等人(2002)《细菌学杂志》184(15):4177-4186,所述文献出于所有目的通过引用整体并入本文)。例如,噬菌体的整合酶基因和(例如,U153或PSA李斯特菌)附着位点可以用于将异源基因插入到相应的附着位点,所述附着位点可以是基因组中的任何适当的位点(例如,comK或arg tRNA基因的3'端)。内源性噬菌体可以在整合异源核酸之前从所利用的附着位点固化。此类方法可以导致例如单拷贝整合体。为了避免“噬菌体固化步骤”,可以使用基于PSA噬菌体的噬菌体整合系统(参见例如,Lauer等人(2002)《细菌学杂志》184:4177-4186,所述文献出于所有目的通过引用整体并入本文)。保持整合的基因可能需要例如通过抗生素进行连续选择。可替代地,可以建立不需要使用抗生素进行选择的基于噬菌体的染色体整合系统。相反,营养缺陷型宿主菌株可以得到补充。例如,可以使用用于临床应用的基于噬菌体的染色体整合系统,其中使用对必需酶(包含例如,D-丙氨酸消旋酶)营养缺陷的宿主菌株(例如,Lm dal(-)dat(-))。
也可以使用缀合将遗传物质和/或质粒引入到细菌中。用于缀合的方法是公知的,并且在以下文献中进行描述:例如,Nikodinovic等人(2006)《质粒(Plasmid)》56(3):223-227以及Auchtung等人(2005)《美国国家科学院院刊》102(35):12554-12559,所述文献中的每个文献出于所有目的通过引用整体并入本文。
在具体实例中,重组细菌或李斯特菌菌株可以包括对重组融合多肽进行编码的核酸,所述重组融合多肽基因组整合到细菌或李斯特菌基因组中,作为具有内源性actA序列(对ActA蛋白进行编码)或内源性hly序列(对LLO蛋白进行编码)的开放阅读框。例如,融合多肽的表达和分泌可以在内源性actA启动子和ActA信号序列的控制下,或者可以在内源性hly启动子和LLO信号序列的控制下。作为另一个实例,对重组融合多肽进行编码的核酸可以替换对ActA蛋白进行编码的actA序列或对LLO蛋白进行编码的hly序列。
重组细菌或李斯特菌菌株的选择可以通过任何方式实现。例如,可以使用抗生素选择。抗生素抗性基因可以用于分子生物学和疫苗制备中常用的常规选择和克隆过程。示例性抗生素抗性基因包含赋予对氨苄青霉素、青霉素、甲氧西林、链霉素、红霉素、卡那霉素、四环素、氯霉素(CAT)、新霉素、潮霉素和庆大霉素的抗性的基因产物。可替代地,可以使用营养缺陷型菌株,并且可以使用外源性代谢基因替代或辅助抗生素抗性基因来进行选择。作为实例,为了选择包括对本文所提供的代谢酶或补充基因进行编码的质粒的营养缺陷型细菌,可以在将选择表达对代谢酶进行编码的基因(例如,氨基酸代谢基因)或补充基因的培养基中生长经转化的营养缺陷型细菌。可替代地,温度敏感性质粒可以用于选择重组体或用于选择重组体的任何其它已知方式。
B.细菌或李斯特菌菌株的减毒
本文所公开的重组细菌菌株(例如,重组李斯特菌菌株)可以被减毒。术语“减毒”涵盖细菌引起宿主动物疾病的能力的减弱。例如,尽管减毒的李斯特菌能够在培养中生长和保持,但与野生型李斯特菌相比,减毒的李斯特菌菌株的致病特性可能会降低。在一些实施例中,作为实例,用减毒的李斯特菌对BALB/c小鼠进行静脉内接种,50%的经接种的动物存活时的致死剂量(LD50)比野生型李斯特菌的LD50增加高了至少约10倍、至少约100倍、至少约1,000倍、至少约10,000倍或至少约100,000倍。因此,李斯特菌的减毒菌株是一种不会杀死其所施用的动物的菌株,或者是一种仅当所施用的细菌的数量远远大于杀死同一只动物所需的野生型非减毒细菌的数量时才会杀死动物的菌株。减毒细菌也应解释为意指在一般环境中不能够复制的细菌,因为所述一般环境中不存在细菌生长所需的营养。因此,细菌限于在提供所需营养的受控环境中复制。减毒菌株对于环境是安全的,因为其无法不受控地复制
(1)使细菌和李斯特菌菌株减毒的方法
减毒可以通过任何已知的方式来完成。例如,此类减毒菌株可能缺乏一种或多种内源性毒力基因或一种或多种内源性代谢基因。本文公开了此类基因的实例,并且可以通过使本文所公开的基因中的任何一种基因或任何组合失活来实现减毒。失活可以例如通过缺失或通过突变(例如,失活突变)来实现。术语“突变”包含对序列(核酸或氨基酸序列)的任何类型的突变或修饰,并且可以涵盖缺失、截短、插入、取代、破坏或转位。例如,突变可以包含移码突变、引起蛋白质提前终止的突变或影响基因表达的调节序列的突变。诱变可以使用重组DNA技术或使用诱变化学品或辐射的传统诱变技术,并且随后选择突变体来完成。缺失突变体可以是优选的,因为伴随着逆转的可能性低。术语“代谢基因”是指对酶进行编码的基因,所述酶参与宿主细菌利用或需要的营养素的合成或对所述合成必不可少。例如,酶可以参与宿主细菌的持续生长所需的营养素的合成或对所述合成必不可少。术语“毒力”基因包含这样一种基因,其在生物体基因组中的存在或活性有助于生物体的致病性(例如,使生物体能够在宿主中实现生态位的定殖(包含附着到细胞))、免疫逃避(逃避宿主的免疫应答)、免疫抑制(抑制宿主的免疫应答)、进入或退出细胞或从宿主体内获取营养素。
此类减毒菌株的具体实例是单核细胞增生性李斯特菌(Lm)dal(-)dat(-)(Lmdd)。此类减毒菌株的另一个实例是Lm dal(-)dat(-)ΔactA(LmddA)。参见例如,US 2011/0142791,所述文献出于所有目的通过引用整体并入本文。LmddA基于由于内源性毒力基因actA的缺失而减毒的李斯特菌菌株。此类菌株可以通过dal基因的互补而保留用于体内和体外抗原表达的质粒。可替代地,LmddA可以是在内源性dal、dat和actA基因中具有突变的dal/dat/actA李斯特菌。此类突变可以是例如缺失或其它失活突变。
减毒菌株的另一个具体实例是Lm prfA(-)或在prfA基因中具有部分缺失或失活突变的菌株。PrfA蛋白控制调节子的表达,所述调节子包括Lm定殖其脊椎动物宿主所需的必需毒力基因;因此,prfA突变会大大削弱PrfA活化PrfA依赖性的毒力基因的表达的能力。
减毒菌株的又另一个具体实例是Lm inlB(-)actA(-),其中缺失了对细菌自然毒力至关重要的两种基因—内化蛋白B和act A。
减毒的细菌或李斯特菌菌株的其它实例包含缺乏一种或多种内源性毒力基因的细菌或李斯特菌菌株。此类基因的实例包含李斯特菌中的actA、prfA、plcB、plcA、inlA、inlB、inlC、inlJ和bsh。减毒的李斯特菌菌株也可以是上述菌株中的任何菌株的双突变体或三突变体。减毒的李斯特菌菌株可以包括基因中的每个基因的突变或缺失,或包括例如本文所提供的基因(例如,包含actA、prfA和dal/dat基因)中的任何基因的多达十个基因的突变或缺失。例如,减毒的李斯特菌菌株可以包括内源性内化蛋白C(inlC)基因的突变或缺失和/或内源性actA基因的突变或缺失。可替代地,减毒的李斯特菌菌株可以包括内源性内化蛋白B(inlB)基因的突变或缺失和/或内源性actA基因的突变或缺失。可替代地,减毒的李斯特菌菌株可以包括内源性inlB、inlC和actA基因的突变或缺失。参与此过程的内源性actA基因和/或内源性inlC基因或内源性inlB基因的缺失抑制了李斯特菌向邻近细胞的转位,从而导致高水平的减毒,增强了免疫原性,并且可用作菌株主链。减毒的李斯特菌菌株也可以是包括plcA和plcB两者的突变或缺失的双突变体。在一些情况下,可以从EGD李斯特菌主链构建菌株。
细菌或李斯特菌菌株也可以是在代谢基因中具有突变的营养缺陷型菌株。作为一个实例,菌株可以缺乏一种或多种内源性氨基酸代谢基因。例如,缺乏D-丙氨酸的李斯特菌的营养缺陷型菌株的产生可以通过多种公知的方式来完成,包含缺失突变、插入突变、移码突变、导致蛋白质提前终止的突变或影响基因表达的调节序列的突变。缺失突变体可以是优选的,因为伴随着营养缺陷型表型的逆转的可能性低。例如,可以在简单的实验室培养测定中测试根据本文提出的方案产生的D-丙氨酸的突变体在不存在D-丙氨酸的情况下生长的能力。可以选择在不存在此化合物的情况下无法生长的突变体。
内源性氨基酸代谢基因的实例包含维生素合成基因、对泛酸合酶进行编码的基因、D-谷氨酸合酶基因、D-丙氨酸氨基转移酶(dat)基因、D-丙氨酸消旋酶(dal)基因、dga、参与合成二氨基庚二酸(DAP)的基因、参与合成半胱氨酸合酶A(cysK)的基因、维生素B12非依赖性蛋氨酸合酶、trpA、trpB、trpE、asnB、gltD、gltB、leuA、argG和thrC。李斯特菌菌株可能缺乏两个或更多个此类基因(例如,dat和dal)。D-谷氨酸的合成部分地受dal基因的控制,所述基因参与D-glu+pyr向α-酮戊二酸+D-ala的转化以及逆反应。
作为另一个实例,减毒的李斯特菌菌株可能缺乏内源性合酶基因,如氨基酸合成基因。此类基因的实例包含folP、对二氢尿苷合酶家族蛋白进行编码的基因、ispD、ispF、对磷酸烯醇丙酮酸合酶进行编码的基因、hisF、hisH、fliI、对核糖体大亚基假尿苷合酶进行编码的基因、ispD、对双功能GMP合酶/谷氨酰胺酰胺转移酶蛋白进行编码的基因、cobS、cobB、cbiD、对尿卟啉-III C-甲基转移酶/尿卟啉原-III合酶进行编码的基因、cobQ、uppS、truB、dxs、mvaS、dapA、ispG、folC、对柠檬酸合酶进行编码的基因、argJ、对3-脱氧-7-磷酸庚糖酮酸合酶进行编码的基因、对吲哚-3-甘油磷酸合酶进行编码的基因、对邻氨基苯甲酸合酶/谷氨酰胺酰胺转移酶组分进行编码的基因、menB、对甲基萘醌特异性异分支酸合酶进行编码的基因、对磷酸核糖基甲酰基甘氨酰胺合酶I或II进行编码的基因、对磷酸核糖基氨基咪唑-琥珀羧胺合酶进行编码的基因、carB、carA、thyA、mgsA、aroB、hepB、rluB、ilvB、ilvN、alsS、fabF、fabH、对假尿苷合酶进行编码的基因、pyrG、truA、pabB以及atp合酶基因(例如,atpC、atpD-2、aptG、atpA-2等)。
减毒的李斯特菌菌株可能缺乏内源性phoP、aroA、aroC、aroD或plcB。作为又另一个实例,减毒的李斯特菌菌株可能缺乏内源性肽转运体。实例包含对以下各项进行编码的基因:ABC转运体/ATP结合/渗透酶蛋白、寡肽ABC转运体/寡肽结合蛋白、寡肽ABC转运体/渗透酶蛋白、锌ABC转运体/锌结合蛋白、糖ABC转运体、磷酸盐转运体、ZIP锌转运体、EmrB/QacA家族的耐药性转运体、硫酸盐转运体、质子依赖性寡肽转运体、镁转运体、甲酸盐/亚硝酸盐转运体、亚精胺/尿素ABC转运体、Na/Pi-共转运体、磷酸糖转运体、谷氨酰胺ABC转运体、主要促进剂家族转运体、甘氨酸甜菜碱/L-脯氨酸ABC转运体、钼ABC转运体、磷壁酸ABC转运体、钴ABC转运体、铵转运体、氨基酸ABC转运体、细胞分裂ABC转运体、锰ABC转运体、铁化合物ABC转运体、麦芽糖/麦芽糊精ABC转运体、Bcr/CflA家族的耐药性转运体以及上述蛋白质之一的亚基。
其它减毒的细菌和李斯特菌菌株可能缺乏内源性代谢酶,所述酶代谢用于细菌生长过程、复制过程、细胞壁合成、蛋白质合成、脂肪酸代谢或任何其它生长或复制过程的氨基酸。同样,减毒的菌株可能缺乏内源性代谢酶,所述酶可以催化细胞壁合成中所用的氨基酸的形成、可以催化细胞壁合成中所用的氨基酸的合成或者可以参与细胞壁合成中所用的氨基酸的合成。可替代地,氨基酸可以用于细胞壁生物发生。可替代地,代谢酶是细胞壁组分D-谷氨酸的合成酶。
其它减毒的李斯特菌菌株可能缺乏由D-谷氨酸合成基因、dga、alr(丙氨酸消旋酶)基因或任何其它参与丙氨酸合成的酶编码的代谢酶。李斯特菌菌株可能缺乏的代谢酶的又其它实例包含由以下各项编码的酶:serC(磷酸丝氨酸氨基转移酶)、asd(天冬氨酸β-半醛脱氢酶;参与细胞壁成分二氨基庚二酸的合成)、对gsaB-谷氨酸-1-半醛氨基转移酶(催化由(S)-4-氨基-5-氧代戊酸形成5-氨基乙酰丙酸酯)进行编码的基因、hemL(催化由(S)-4-氨基-5-氧代戊酸形成5-氨基乙酰丙酸酯)、aspB(催化由L-天冬氨酸和2-氧代戊二酸形成草酰乙酸和L-谷氨酸的天冬氨酸氨基转移酶)、argF-1(参与精氨酸的生物合成)、aroE(参与氨基酸的生物合成)、aroB(参与3-脱氢奎宁酸酯的生物合成)、aroD(参与氨基酸的生物合成)、aroC(参与氨基酸的生物合成)、hisB(参与组氨酸的生物合成)、hisD(参与组氨酸的生物合成)、hisG(参与组氨酸的生物合成)、metX(参与蛋氨酸的生物合成)、proB(参与脯氨酸的生物合成)、argR(参与精氨酸的生物合成)、argJ(参与精氨酸的生物合成)、thil(参与硫胺素的生物合成)、LMOf2365_1652(参与色氨酸的生物合成)、aroA(参与色氨酸的生物合成)、ilvD(参与缬氨酸和异亮氨酸的生物合成)、ilvC(参与缬氨酸和异亮氨酸的生物合成)、leuA(参与参与亮氨酸的生物合成)、dapF(参与赖氨酸的生物合成)和thrB(苏氨酸的生物合成)(所有基因库登录号NC_002973)。
减毒的李斯特菌菌株可以通过其它代谢酶(如tRNA合成酶)的突变产生。例如,代谢酶可以由trpS基因编码,所述基因对色氨酰基tRNA合成酶进行编码。例如,宿主菌株细菌可以是Δ(trpS aroA),并且两种标志物都可以包含在整合载体中。
可以被突变以产生减毒的李斯特菌菌株的代谢酶的其它实例包含由以下各项进行编码的酶:murE(参与二氨基庚二酸的合成;基因库登录号:NC_003485)、LMOf2365_2494(参与磷壁酸的生物合成)、WecE(脂多糖生物合成蛋白rffA;基因库登录号:AE014075.1)或amiA(N-乙酰基胞壁酰-L-丙氨酸酰胺酶)。代谢酶的又其它实例包含天冬氨酸氨基转移酶、组氨丁醇磷酸氨基转移酶(基因库登录号NP_466347)或细胞壁磷壁酸糖基化蛋白GtcA。
可以被突变以产生减毒的李斯特菌菌株的代谢酶的其它实例包含用于肽聚糖组分或前体的合成酶。组分可以是例如UDP-N-乙酰基胞壁酰五肽、UDP-N-乙酰基葡糖胺、MurNAc-(五肽)-焦磷酰基-十一异戊烯、GlcNAc-p-(1,4)-MurNAc-(五肽)-焦磷酰基十一异戊烯或任何其它肽聚糖组分或前体。
可以被突变以产生减毒的李斯特菌菌株的代谢酶的其它实例包含由murG、murD、murA-1或murA-2(均在基因库登录号NC_002973中列出)编码的代谢酶。可替代地,代谢酶可以是肽聚糖组分或前体的任何其它合成酶。代谢酶也可以是转糖基酶、转肽酶、羧肽酶、任何其它种类的代谢酶或任何其它代谢酶。例如,代谢酶可以是任何其它李斯特菌代谢酶或任何其它单核细胞增生性李斯特菌代谢酶。
如上文针对李斯特菌所描述的,可以通过突变其它细菌菌株中的对应直系同源基因来使其它细菌菌株减毒。
(2)补充减毒的细菌和李斯特菌菌株的方法
本文所公开的减毒的细菌或李斯特菌菌株可以进一步包括以下核酸,所述核酸包括补充基因或对补充减毒突变(例如,补充营养缺陷型李斯特菌菌株的营养缺陷)的代谢酶进行编码。例如,具有对如本文所公开的融合多肽进行编码的第一开放阅读框的核酸可以进一步包括第二开放阅读框,所述第二开放阅读框包括补充基因或对补充代谢酶进行编码。可替代地,第一核酸可以对融合多肽进行编码,并且单独的第二核酸可以包括补充基因或对补充代谢酶进行编码。
补充基因可以是染色体外的,或者可以整合到细菌或李斯特菌基因组中。例如,营养缺陷型李斯特菌菌株可以包括附加型质粒,所述附加型质粒包括对代谢酶进行编码的核酸。此类质粒将以附加或染色体外的方式包含在李斯特菌中。可替代地,营养缺陷型李斯特菌菌株可以包括整合型质粒(即,整合载体),所述整合型质粒包括对代谢酶进行编码的核酸。此类整合型质粒可以用于整合到李斯特菌染色体中。优选地,附加型质粒或整合型质粒缺乏抗生素抗性标志物。
可以使用代谢基因替代或辅助抗生素抗性基因来进行选择。作为实例,为了选择包括对本文所提供的代谢酶或补充基因进行编码的质粒的营养缺陷型细菌,可以在将选择表达对代谢酶进行编码的基因(例如,氨基酸代谢基因)或补充基因的培养基中生长经转化的营养缺陷型细菌。例如,可以用包括用于D-谷氨酸合成的基因的质粒转化用于D-谷氨酸合成的营养缺陷型细菌,并且营养缺陷型细菌将在不存在D-谷氨酸的情况下生长,而没有用质粒转化的营养缺陷型细菌或不表达对用于D-谷氨酸合成的蛋白质进行编码的质粒的营养缺陷型细菌将不会生长。类似地,当转化并表达包括对用于D-丙氨酸合成的氨基酸代谢酶进行编码的核酸的质粒时,用于D-丙氨酸合成的营养缺陷型细菌将在不存在D-丙氨酸的情况下生长。用于制备包括或缺乏必需的生长因子、补充剂、氨基酸、维生素、抗生素等的适当培养基的此类方法是公知的,并且可商购获得。
一旦在适当的培养基中选择了包括对本文所提供的代谢酶或补充基因进行编码的质粒的营养缺陷型细菌,就可以在存在选择压力的情况下繁殖细菌。此类繁殖可以包括使细菌在没有营养缺陷型因子的培养基中生长。在营养缺陷型细菌中表达代谢酶或补充基因的质粒的存在确保了质粒将与细菌一起复制,从而不断地选择具有所述质粒的细菌。通过调节包括质粒的营养缺陷型细菌所生长的培养基的体积,可以容易地扩大细菌或李斯特菌菌株的产生。
在一个具体实例中,减毒的菌株是具有dal和dat的缺失或dal和dat中的失活突变的菌株(例如,单核细胞增生性李斯特菌(Lm)dal(-)dat(-)(Lmdd)或Lm dal(-)dat(-)ΔactA(LmddA)),并且补充基因编码丙氨酸消旋酶(例如,由dal基因编码)或D-氨基酸氨基转移酶(例如,由dat基因编码)。示例性丙氨酸消旋酶蛋白可以具有SEQ ID NO:76中所示的序列(由SEQ ID NO:78编码;基因库登录号:AF038438),或者可以是SEQ ID NO:76的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段或同工型的片段。丙氨酸消旋酶蛋白也可以是任何其它李斯特菌丙氨酸消旋酶蛋白。可替代地,丙氨酸消旋酶蛋白可以是任何其它革兰氏阳性丙氨酸消旋酶蛋白或任何其它丙氨酸消旋酶蛋白。示例性D-氨基酸氨基转移酶蛋白可以具有SEQ ID NO:77中所示的序列(由SEQ ID NO:79编码;基因库登录号:AF038439),或者可以是SEQ ID NO:77的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段或同工型的片段。D-氨基酸氨基转移酶蛋白也可以是任何其它李斯特菌D-氨基酸氨基转移酶蛋白。可替代地,D-氨基酸氨基转移酶蛋白可以是任何其它革兰氏阳性D-氨基酸氨基转移酶蛋白或任何其它D-氨基酸氨基转移酶蛋白。
在另一个具体实例中,减毒的菌株是具有prfA的缺失或prfA中的失活突变的菌株(例如,Lm prfA(-)),并且补充基因对PrfA蛋白进行编码。例如,补充基因可以对恢复部分PrfA功能的突变PrfA(D133V)蛋白进行编码。野生型PrfA蛋白的实例在SEQ ID NO:80中示出(由SEQ ID NO:81中所示的核酸编码),并且D133V突变PrfA蛋白的实例在SEQ ID NO:82中示出(由SEQ ID NO:83中所示的核酸编码)。补充PrfA蛋白可以是SEQ ID NO:80或82的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段或同工型的片段。PrfA蛋白也可以是任何其它李斯特菌PrfA蛋白。可替代地,PrfA蛋白可以是任何其它革兰氏阳性PrfA蛋白或任何其它PrfA蛋白。
在另一个实例中,细菌菌株或李斯特菌菌株可以包括actA基因的缺失或actA基因中的失活突变,并且补充基因可以包括actA基因以补充突变并且恢复对李斯特菌菌株的功能。
其它营养缺陷型菌株和互补系统也可以与本文所提供的方法和组合物一起使用。
IV.重组融合多肽
本文所公开的重组细菌或李斯特菌菌株中的重组融合多肽可以呈任何形式。一些此类融合多肽可以包括与一种或多种疾病相关型抗原肽融合的含PEST的肽。其它此类重组融合多肽可以包括一种或多种疾病相关型抗原肽,并且其中所述融合多肽不包括含PEST的肽。
重组融合多肽的另一个实例包括从N端到C端的细菌分泌序列、泛素(Ub)蛋白和一种或多种疾病相关型抗原肽(即,串联的,如Ub-肽1-肽2)。可替代地,如果使用两种或更多种疾病相关型抗原肽,则可以使用单独的融合多肽的组合,其中每个抗原肽都与其自身的分泌序列和Ub蛋白(例如,Ub1-肽1;Ub2-肽2)融合。
还公开了对此类重组融合多肽进行编码的核酸(被称为小基因构建体)。此类小基因核酸构建体可以进一步包括通过每个开放阅读框之间的Shine-Dalgarno核糖体结合位点核酸序列连接的两个或更多个开放阅读框。例如,小基因核酸构建体可以进一步包括通过每个开放阅读框之间的Shine-Dalgarno核糖体结合位点核酸序列连接的两个到四个开放阅读框。每个开放阅读框可以编码不同的多肽。在一些核酸构建体中,对融合多肽的羧基端进行编码的密码子后面是两个终止密码子,以确保蛋白质合成的终止。
细菌信号序列可以是李斯特菌信号序列,如Hly或ActA信号序列,或任何其它已知的信号序列。在其它情况下,信号序列可以是LLO信号序列。示例性LLO信号序列在SEQ IDNO:97中示出。信号序列可以是细菌的,可以是宿主细菌的天然序列(例如,单核细胞增生性李斯特菌,如secA1信号肽)或者可以是宿主细菌的外源序列。信号肽的具体实例包含来自乳酸乳球菌(Lactococcus lactis)的Usp45信号肽、来自炭疽芽孢杆菌(Bacillusanthracis)的保护性抗原信号肽、来自单核细胞增生性李斯特菌的secA2信号肽(如p60信号肽)以及Tat信号肽(如枯草芽孢杆菌(B.subtilis)Tat信号肽)(例如,PhoD)。在具体实例中,分泌信号序列来自李斯特菌蛋白,如ActA300分泌信号或ActA100分泌信号。示例性ActA信号序列在SEQ ID NO:98中示出。
泛素可以是例如全长蛋白。在进入宿主细胞的细胞溶质时,通过水解酶的作用,从本文所提供的核酸构建体中表达的泛素可以在羧基端与从所述核酸构建体中表达的其余重组融合多肽切割开。这释放了融合多肽的氨基端,从而在宿主细胞的细胞溶质中产生了肽。
融合多肽内的抗原肽的选择、变异和排列在本文其它地方详细讨论,并且疾病相关型抗原肽的实例在本文其它地方更详细地讨论。
重组融合多肽可以包括一个或多个标签。例如,重组融合多肽可以包括一个或多个抗原肽的N端和/或C端的一个或多个肽标签。标签可以直接与抗原肽融合或通过接头与抗原肽连接(所述接头的实例在本文其它地方公开)。标签的实例包含以下各项:FLAG标签;2xFLAG标签;3xFLAG标签;His标签,6xHis标签;以及SIINFEKL标签。示例性SIINFEKL标签在SEQ ID NO:16中示出(由SEQ ID NO:1-15中所示的核酸中的任何一个核酸编码)。示例性3xFLAG标签在SEQ ID NO:32中示出(由SEQ ID NO:17-31中所示的核酸中的任何一个核酸编码)。示例性变体3xFLAG标签在SEQ ID NO:99中示出。可以一起使用两个或更多个标签,如2xFLAG标签和SIINFEKL标签、3xFLAG标签和SIINFEKL标签或6xHis标签和SIINFEKL标签。如果使用两个或更多个标签,则所述标签可以以任何顺序定位在重组融合多肽内的任何位置。例如,两个标签可以位于重组融合多肽的C端,两个标签可以位于重组融合多肽的N端,两个标签可以定位在重组融合多肽的内部,一个标签可以位于重组融合多肽的C端并且一个标签位于重组融合多肽的N端,一个标签可以位于重组融合多肽的C端并且一个标签在重组融合多肽内部,或者一个标签可以位于重组融合多肽的N端并且一个标签在重组融合多肽内部。其它标签包含几丁质结合蛋白(CBP)、麦芽糖结合蛋白(MBP)、谷胱甘肽-S-转移酶(GST)、硫氧还蛋白(TRX)和聚(NANP)。特定重组融合多肽包括C端SIINFEKL标签。此类标签可以允许容易地检测重组融合蛋白,确认重组融合蛋白的分泌,或通过追踪对这些“标签”序列肽的免疫应答来追踪分泌性融合多肽的免疫原性。可以使用多种试剂来监测这种免疫应答,所述试剂包含例如对这些标签具有特异性的单克隆抗体和DNA或RNA探针。
本文所公开的重组融合多肽可以由重组李斯特菌菌株表达或可以从用于蛋白质表达和分离的其它载体和细胞系统中表达和分离。包括表达此类抗原肽的重组李斯特菌菌株可以用于例如包括此类重组李斯特菌的免疫原性组合物中,并且用于包括重组李斯特菌菌株和佐剂的疫苗中。在李斯特菌菌株的宿主细胞系统中和除李斯特菌之外的宿主细胞系统中,一个或多个抗原肽以具有非溶血性截短形式的LLO、ActA或PEST样序列的融合多肽的形式表达,可以增强抗原肽的免疫原性。
还公开了对此类重组融合多肽进行编码的核酸。核酸可以呈任何形式。核酸可以包括DNA或RNA或由DNA或RNA组成,并且可以是单链或双链的。核酸可以呈质粒的形式,如附加型质粒、多拷贝附加型质粒或整合型质粒。可替代地,核酸可以呈病毒载体、噬菌体载体或细菌人工染色体的形式。此类核酸可以具有一个开放阅读框或可以具有两个或更多个开放阅读框(例如,对重组融合多肽进行编码的开放阅读框和对代谢酶进行编码的第二开放阅读框)。在一个实例中,此类核酸可以包括通过每个开放阅读框之间的Shine-Dalgarno核糖体结合位点核酸序列连接的两个或更多个开放阅读框。例如,核酸可以包括通过每个开放阅读框之间的Shine-Dalgarno核糖体结合位点核酸序列连接的两个到四个开放阅读框。每个开放阅读框可以编码不同的多肽。在一些核酸中,对融合多肽的羧基端进行编码的密码子后面是两个终止密码子,以确保蛋白质合成的终止。
A.抗原肽
疾病相关型肽包含来自在特定疾病中表达的蛋白质的肽。例如,此类肽可以来自在疾病组织中表达但不在对应的正常组织中表达的蛋白质,或在疾病组织中异常高水平表达的蛋白质。如本文所使用的,术语“疾病”旨在通常是同义词,并且与术语“病症”和“病状”(如医学病状)可互换地使用,因为它们均反映出人体或动物体的异常情况或其部分中的损害正常功能的一部分的异常情况,通常表现为明显的体征和症状,并且导致人体或动物的生命持续时间或生活质量降低。疾病相关型抗原肽的实例可以包含人乳头瘤病毒(HPV)E7或E6、前列腺特异性抗原(PSA)、嵌合Her2抗原、Her2/neu嵌合抗原。人乳头瘤病毒可以是HPV 16或HPV 18。抗原肽还可以包含可操作地串联连接的HPV16E6、HPV16 E7、HPV18 E6、HPV18 E7抗原或可操作地串联连接至HPV抗原肽的HPV16抗原肽。
融合多肽可以包含单个抗原肽,或者可以包含两个或更多个抗原肽。每个抗原肽可以具有足以诱导免疫应答的任何长度,并且每个抗原肽可以具有相同的长度,或者抗原肽可以具有不同的长度。例如,本文所公开的抗原肽的长度可以是5-100个、15-50个或21-27个氨基酸,或者长度可以是15-100个、15-95个、15-90个、15-85个、15-80个、15-75个、15-70个、15-65个、15-60个、15-55个、15-50个、15-45个、15-40个、15-35个、15-30个、20-100个、20-95个、20-90个、20-85个、20-80个、20-75个、20-70个、20-65个、20-60个、20-55个、20-50个、20-45个、20-40个、20-35个、20-30个、11-21个、15-21个、21-31个、31-41个、41-51个、51-61个、61-71个、71-81个、81-91个、91-101个、101-121个、121-141个、141-161个、161-181个、181-201个、8-27个、10-30个、10-40个、15-30个、15-40个、15-25个、1-10个、10-20个、20-30个、30-40个、1-100个、5-75个、5-50个、5-40个、5-30个、5-20个、5-15个、5-10个、1-75个、1-50个、1-40个、1-30个、1-20个、1-15个、1-10个、8-11个或11-16个氨基酸。例如,抗原肽的长度可以为至少15个、16个、17个、18个、19个、20个、21个、22个、23个、24个、25个、26个、27个、28个、29个、30个、31个、32个、33个、34个、35个、36个、37个、38个、39个、40个、41个、42个、43个、44个、45个、46个、47个、48个、49个、50个、51个、52个、53个、54个、55个、56个、57个、58个、59或60个氨基酸。抗原肽的一些具体实例是长度为21或27个氨基酸。其它抗原肽可以是全长蛋白或其片段。
作为一个实例,抗原肽可以包括新表位。这些新表位可以是例如患者特异性(即,受试者特异性)癌症突变。可以在用于产生个性化免疫疗法的过程中产生包括新表位的抗原肽,所述过程包括将从受试者的癌症样品中提取的核酸与从正常或健康参考样品中提取的核酸进行比较,以鉴别与正常或健康样品相比,癌症样品中存在的体细胞突变或序列差异。例如,这些突变或序列差异可以是体细胞的非同义的错义突变或体细胞的移码突变,并且可以编码经表达的氨基酸序列。表达此类体细胞突变或序列差异的肽可以被称为“新表位”。癌症特异性新表位可以指在参考样品(如正常的非癌性或种系细胞或组织)中不存在但在癌症样品中发现的表位。例如,这包含在正常的非癌性或种系细胞中找到对应表位的情况,但由于癌细胞中的一种或多种突变,表位的序列发生改变,从而导致新表位。新表位可以包括突变的表位,并且可以在突变的任一侧或两侧都包括非突变的序列。
作为另一个实例,抗原肽可以包括复发性癌症突变。例如,本文所公开的重组融合多肽可以包括与两个或更多个抗原肽融合的含PEST的肽(即,串联的,如PEST-肽1-肽2),或者可以包括不与含PEST的肽融合的两个或更多个抗原肽,其中每个抗原肽都包括单一的复发性癌症突变(即,蛋白质的氨基酸序列或由基因中的单一、不同、非同义的复发性癌症突变编码的序列中的单一复发性变化),并且其中所述抗原肽中的至少两个抗原肽包括不同的复发性癌症突变,并且是同一癌症相关型蛋白的片段。可替代地,抗原肽中的每个抗原肽都可以包括与不同的癌症相关型蛋白不同的复发性癌症突变。可替代地,可以使用单独的融合多肽的组合,其中每个抗原肽都与其自身的含PEST的肽(例如,PEST1-肽1;PEST2-肽2)融合(或不融合)。任选地,一些或所有片段是同一癌症相关型蛋白的非连续片段。非连续片段是在蛋白质序列中不按顺序出现的片段(例如,第一个片段由残基10-30组成,并且第二个片段由残基100-120组成;或者第一个片段由残基10-30组成,并且第二个片段由残基20-40组成)。任选地,抗原肽中的每个抗原肽都包括与单一类型的癌症不同的复发性癌症突变。
复发性癌症突变可以来自癌症相关型蛋白。术语“癌症相关型蛋白”包含具有突变的蛋白质,所述突变在多种类型的癌症中发生、在患有特定类型的癌症的多个受试者中发生或与一种或多种类型的癌症的发生或发展相关。例如,癌症相关型蛋白可以是致癌蛋白(即,具有可能有助于癌症进展的活性的蛋白质,如调节细胞生长的蛋白质),或者其可以是肿瘤抑制蛋白(即,通常用于如通过细胞周期的负调节或通过促进细胞凋亡来减轻癌症形成的可能性的蛋白质)。优选地,癌症相关型蛋白具有“突变热点”。突变热点是蛋白质编码基因中的氨基酸位置,其突变频率比不存在选择时预期的突变频率更高(优选地通过体细胞取代而不是其它体细胞异常,如转位、扩增和缺失)。此类热点突变可以发生在多种类型的癌症中和/或可以在多个癌症患者之间共享。突变热点表明跨肿瘤样品群体的选择性压力。肿瘤基因组含有通过影响基因(即,肿瘤驱动基因)来“驱动”肿瘤发生的复发性癌症突变,所述基因在发生更改时赋予肿瘤细胞选择性生长优势。此类肿瘤驱动基因可以通过以下进行鉴别:例如通过根据背景突变速率(即,复发性)鉴别突变频率比预期更高的基因;通过鉴别跨肿瘤样品表现出其它阳性选择信号的基因(例如,与沉默突变相比,非沉默突变的发生率高,或者倾向于功能性突变的积累);通过基于以下知识来利用维持蛋白质序列的某些区域中的突变的趋势,即:失活突变沿蛋白质的序列分布,而功能获得性突变则倾向于具体在特定残基或结构域中发生;或者通过利用具体功能性残基(如磷酸化位点)中突变的过量表达。这些突变中的许多突变经常发生在生物活性蛋白的功能性区域(例如,激酶结构域或结合结构域)或中断活性位点(例如,磷酸化位点),从而导致功能丧失或功能获得性突变,或者所述突变可以以使得蛋白质的三维结构和/或电荷平衡受到足够扰乱从而干扰正常功能的方式发生。对大量肿瘤的基因组分析表明,突变通常发生在有限数量的氨基酸定位。因此,大多数常见突变可以由相对少量的潜在肿瘤相关性抗原或T细胞表位代表。
“复发性癌症突变”是在多种类型的癌症和/或在患有特定类型的癌症的多个受试者中发生的蛋白质的氨基酸序列的变化。与癌症相关的此类突变可以导致通常在对应的健康组织中不存在的肿瘤相关性抗原。
具有跨多种癌症或在多种癌症患者之间发生的具有共同突变的肿瘤驱动基因和癌症相关型蛋白是已知的,并且存在跨多个肿瘤样品和多种肿瘤类型的测序数据。参见例如,Chang等人(2016)《自然-生物技术(Nat Biotechnol)》34(2):155-163;Tamborero等人(2013)《科学报告(Sci Rep)》3:2650,所述文献中的每个文献通过引用整体并入本文。
每个抗原肽也可以是亲水性的,或者可以评分达到或低于某个亲水性阈值,这可以预测单核细胞增生性李斯特菌或另一种所关注的细菌的分泌性。例如,可以通过Kyte和Doolittle亲水性指数21个氨基酸窗口对抗原肽进行评分,并且可以排除所有高于临界值(约1.6)的评分,因为它们不太可能可由单核细胞增生性李斯特菌分泌。同样,抗原肽的组合或融合多肽可以是亲水性的,或者可以评分达到或低于某个亲水性阈值,这可以预测单核细胞增生性李斯特菌或另一种所关注的细菌的分泌性。
抗原肽可以以任何方式连接在一起。例如,抗原肽可以彼此直接融合而没有插入序列。可替代地,抗原肽可以通过一个或多个接头(如肽接头)间接地彼此连接。在一些情况下,一些成对的相邻抗原肽可以彼此直接融合,而其它成对的抗原肽则可以通过一个或多个接头间接地彼此连接。可以在每对相邻抗原肽之间使用相同的接头,或者可以在不同对相邻抗原肽之间使用任何数量的不同接头。另外,可以在一对相邻抗原肽之间使用一个接头,或者可以在一对相邻抗原肽之间使用多个接头。
任何合适的序列均可以用于肽接头。作为实例,接头序列的长度可以是例如1到约50个氨基酸。一些接头可以是亲水性的。接头可以用于各种目的。例如,接头可以用于增加细菌分泌、促进抗原加工、增加融合多肽的柔韧性、增加融合多肽的刚性或任何其它目的。在一些情况下,不同的氨基酸接头序列分布在抗原肽之间,或者对相同氨基酸接头序列进行编码的不同核酸分布在抗原肽(例如,SEQ ID NO:84-94)之间,以使重复序列最小化。这也可以用于减少二级结构,从而允许在Lm重组载体菌株群体内有效转录、翻译、分泌、保持或稳定对融合多肽进行编码的核酸(例如,质粒)。可以例如基于以下因素中的一个或多个因素选择其它合适的肽接头序列:(1)其能够采用灵活的扩展构象;(2)其不能采用可以与抗原肽上的功能性表位相互作用的二级结构;以及(3)缺乏可以与功能性表位反应的疏水残基或带电残基。例如,肽接头序列可以含有Gly、Asn和Ser残基。还可以在接头序列中使用其它接近中性的氨基酸,如Thr和Ala。可以有效地用作接头的氨基酸序列包含以下文献中公开的氨基酸序列:Maratea等人(1985)《基因(Gene)》40:39-46;Murphy等人,(1986)《美国国家科学院院刊》83:8258-8262;US 4,935,233;以及US 4,751,180,所述文献中的每个文献出于所有目的通过引用整体并入本文。接头的具体实例包含表2中的接头(所述接头中的每个接头都可以单独用作接头,在包括序列的重复序列的接头中或在进一步包括表中的其它序列中的一个或多个序列的接头中),但是也可以设想其它接头(参见例如,ReddyChichili等人(2013)《蛋白质科学(Protein Science)》22:153-167,所述文献出于所有目的通过引用整体并入本文)。除非另有说明,否则“n”表示列出的接头中的重复序列的不确定数量。
表2.接头。
| 肽接头 | 实例 | SEQ ID NO: | 假设目的 |
| (GAS)<sub>n</sub> | GASGAS | 33 | 柔性 |
| (GSA)<sub>n</sub> | GSAGSA | 34 | 柔性 |
| (G)<sub>n</sub>;n=4-8 | GGGG | 35 | 柔性 |
| (GGGGS)<sub>n</sub>;n=1-3 | GGGGS | 36 | 柔性 |
| VGKGGSGG | VGKGGSGG | 37 | 柔性 |
| (PAPAP)<sub>n</sub> | PAPAP | 38 | 刚性 |
| (EAAAK)<sub>n</sub>;n=1-3 | EAAAK | 39 | 刚性 |
| (AYL)<sub>n</sub> | AYLAYL | 40 | 抗原加工 |
| (LRA)<sub>n</sub> | LRALRA | 41 | 抗原加工 |
| (RLRA)<sub>n</sub> | RLRA | 42 | 抗原加工 |
B.含PEST的肽
本文所公开的重组融合蛋白包括含PEST的肽。含PEST的肽可以位于融合多肽的氨基端(N端)末端(即,抗原肽的N端),可以位于融合多肽的羧基端(C端)末端(即,抗原肽的C端),或者可以嵌入在抗原肽内。在一些重组李斯特菌菌株和方法中,含PEST的肽不是融合多肽的一部分,并且与融合多肽分离。抗原肽与PEST样序列(如LLO肽)的融合可以增强抗原肽的免疫原性,并且可以增加细胞介导的和抗肿瘤的免疫应答(即,增加细胞介导的和抗肿瘤的免疫)。参见例如,Singh等人(2005)《免疫学杂志》175(6):3663-3673,所述文献出于所有目的通过引用整体并入本文。
含PEST的肽是包括PEST序列或PEST样序列的肽。真核蛋白中的PEST序列早已被鉴别。例如,通常但不总是由含有几个带正电荷的氨基酸的簇侧接的含有富含脯氨酸(P)、谷氨酸(E)、丝氨酸(S)和苏氨酸(T)(PEST)的氨基酸序列的蛋白质具有快速的细胞内半衰期(Rogers等人(1986)《科学》234:364-369,所述文献出于所有目的通过通过引用整体并入本文)。此外,据报道,这些序列将蛋白质靶向用于降解的泛素-蛋白酶体途径(Rechsteiner和Rogers(1996)《生物化学科学趋势(Trends Biochem.Sci.)》21:267-271,所述文献出于所有目的通过引用整体并入本文)。真核细胞也使用此途径来产生与MHC I类结合的免疫原性肽,并且据假设,PEST序列在产生免疫原性肽的真核蛋白中是丰富的(Realini等人(1994)《欧洲生化学会联合会快报(FEBS Lett.)》348:109-113,所述文献出于所有目的通过引用整体并入本文)。原核蛋白通常不含PEST序列,因为其不具有此酶促途径。然而,在LLO的氨基端已报道了富含氨基酸脯氨酸(P)、谷氨酸(E)、丝氨酸(S)和苏氨酸(T)的PEST样序列,并且据报道,所述序列对于单核细胞增生性李斯特菌的致病性是必要的(Decatur和Portnoy(2000)《科学》290:992-995,所述文献出于所有目的通过引用整体并入本文)。LLO中的此PEST样序列的存在靶向用于通过宿主细胞的蛋白水解机制进行破坏的蛋白质,使得一旦LLO发挥了功能并且促进了单核细胞增生性李斯特菌从吞噬体或溶酶体液泡中逃逸,其在破坏细胞之前就被破坏。
对PEST和PEST样序列的鉴别是公知的并且在以下文献中进行了描述:例如Rogers等人(1986)《科学》234(4774):364-378以及Rechsteiner和Rogers(1996)《生物化学科学趋势》21:267-271,所述文献中的每个文献出于所有目的通过引用整体并入本文。可以使用PEST查找程序来鉴别PEST或PEST样序列。例如,PEST样序列可以是富含脯氨酸(P)、谷氨酸(E)、丝氨酸(S)和苏氨酸(T)残基的区域。任选地,PEST样序列可以由含有几个带正电荷的氨基酸的一个或多个簇侧接。例如,PEST样序列可以被定义为长度为至少12个氨基酸的亲水性区段,其中脯氨酸(P)、天冬氨酸(D)、谷氨酸(E)、丝氨酸(S)和/或苏氨酸(T)残基的局部浓度较高。在一些情况下,PEST样序列不含带正电荷的氨基酸,即精氨酸(R)、组氨酸(H)和赖氨酸(K)。一些PEST样序列可以含有一个或多个内部磷酸化位点,并且这些位点的磷酸化先于蛋白质降解。
在一个实例中,PEST样序列适合于Rogers等人所公开的算法。在另一个实例中,PEST样序列适合于Rechsteiner和Rogers所公开的算法。还可以通过对指定蛋白质序列内带正电荷的氨基酸R、H和K进行初始扫描来鉴别PEST样序列。计算带正电的侧翼之间的所有氨基酸,并且仅进一步考虑氨基酸数量等于或大于窗口大小参数的基序。任选地,PEST样序列必须含有至少一个P、至少一个D或E以及至少一个S或T。
可以通过基于关键氨基酸的局部富集以及基序疏水性的评分参数来完善PEST基序的质量。D、E、P、S和T的富集以质量百分比(w/w)表示,并且针对一当量的D或E、一当量的P和一当量的S或T进行校正。疏水性的计算原则上也可以遵循以下方法:Kyte和Doolittle(1982)《分子生物学杂志(J.Mol.Biol.)》157:105,所述文献出于所有目的通过引用整体并入本文。为了简化计算,使用以下线性转换将Kyte-Doolittle亲水性指数(最初范围从精氨酸的-4.5到异亮氨酸的+4.5)转换为正整数,其得出的值从精氨酸的0到异亮氨酸的90:亲水性指数=10*Kyte-Doolittle亲水性指数+45。
潜在的PEST基序的疏水性也可以计算为每种氨基酸物种的摩尔百分比和疏水性指数乘积的总和。期望的PEST评分是以由下公式表达的以局部富集项和疏水性项的组合的方式得到的:PEST评分=0.55*DEPST-0.5*疏水性指数。
因此,使用上述算法,含PEST的肽可以指评分为至少+5的肽。可替代地,其可以指评分为以下的肽:至少6、至少7、至少8、至少9、至少10、至少11、至少12、至少13、至少14、至少15、至少16、至少17、至少18、至少19、至少20、至少21、至少22、至少23、至少24、至少25、至少26、至少27、至少28、至少29、至少30、至少32、至少35、至少38、至少40或至少45。
任何其它已知的可用方法或算法也可以用于鉴别PEST样序列。参见例如,CaSPredictor(Garay-Malpartida等人(2005)《生物信息学(Bioinformatics)》21增刊1:i169-76,所述文献出于所有目的通过引用整体并入本文)。可以使用的另一种方法是以下方法:通过将氨基酸Ser、Thr、Pro、Glu、Asp、Asn或Gln赋值为1,为每个适当长度的区段(例如30-35个氨基酸的区段)计算PEST指数。PEST残基中的每个残基的系数值(CV)为1,其它AA(非PEST)中的每个的CV为零。
PEST样氨基酸序列的实例是SEQ ID NO:43-51中所示的氨基酸序列。PEST样序列的一个实例是KENSISSMAPPASPPASPKTPIEKKHADEIDK(SEQ ID NO:43)。PEST样序列的另一个实例是KENSISSMAPPASPPASPK(SEQ ID NO:44)。然而,可以使用任何PEST或PEST样氨基酸序列。PEST序列肽是公知的并且在以下文献中进行描述:例如US 7,635,479;US 7,665,238;以及US2014/0186387,所述文献中的每个文献出于所有目的通过引用整体并入本文。
PEST样序列可以来自李斯特菌物种,如来自单核细胞增生性李斯特菌。例如,单核细胞增生性李斯特菌ActA蛋白含有至少四个此类序列(SEQ ID NO:45-48),所述序列中的任何序列均适合用于本文所公开的组合物和方法中。其它类似的PEST样序列包含SEQ IDNO:52-54。链球菌的链球菌溶血素O蛋白也含有PEST序列。例如,化脓性链球菌(Streptococcus pyogenes)的链球菌溶血素O在氨基酸35-51处包括PEST序列KQNTASTETTTTNEQPK(SEQ ID NO:49),并且类马链球菌(Streptococcus equisimilis)的链球菌溶血素O在氨基酸38-54处包括PEST样序列KQNTANTETTTTNEQPK(SEQ ID NO:50)。PEST样序列的另一个实例来自斯氏李斯特菌细胞溶素,由lso基因编码:RSEVTISPAETPESPPATP(例如,SEQ ID NO:51)。
可替代地,PEST样序列可以源自其它原核生物体。其中期望有PEST样氨基酸序列的其它原核生物体包含例如其它李斯特菌物种。
(1)李斯特菌溶血素O(LLO)
可以在本文所公开的组合物和方法中利用的含PEST的肽的一个实例是李斯特菌溶血素O(LLO)肽。LLO蛋白的一个实例是指定为基因库登录号P13128的蛋白质(SEQ ID NO:55;核酸序列在基因库登录号X15127中示出)。SEQ ID NO:55是包含信号序列的前蛋白。前蛋白的前25个氨基酸是信号序列,并且当由细菌分泌时,其会与LLO切割开,从而产生不具有信号序列的504个氨基酸的全长活性LLO蛋白。本文所公开的LLO肽可以包括信号序列或可以包括不包含信号序列的肽。可以使用的示例性LLO蛋白包括以下、基本上由以下组成或由以下组成:SEQ ID NO:55中所示的序列,或SEQ ID NO:55的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段和同工型的片段。可以使用对LLO蛋白的片段或LLO蛋白的同源物、变体、同工型、类似物、同源物的片段、变体的片段或类似物的片段进行编码的任何序列。同源LLO蛋白与参考LLO蛋白的序列同一性可以为例如大于70%、72%、75%、78%、80%、82%、83%、85%、87%、88%、90%、92%、93%、95%、96%、97%、98%或99%。
LLO蛋白的另一个实例在SEQ ID NO:56中示出。可以使用的LLO蛋白可以包括以下、基本上由以下组成或由以下组成:SEQ ID NO:56中所示的序列,或SEQ ID NO:56的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段和同工型的片段。
LLO蛋白的另一个实例是来自单核细胞增生性李斯特菌10403S菌株的LLO蛋白,如基因库登录号:ZP_01942330或EBA21833中所示,或由如基因库登录号:NZ_AARZ01000015或AARZ01000015.1中所示的核酸序列编码。LLO蛋白的另一个实例是来自以下各项的LLO蛋白:单核细胞增生性李斯特菌4b F2365菌株(参见例如,基因库登录号:YP_012823)、EGD-e菌株(参见例如,基因库登录号:NP_463733)或单核细胞增生性李斯特菌的任何其它菌株。LLO蛋白的又另一个实例是来自黄杆菌目细菌HTCC2170(参见例如,基因库登录号:ZP_01106747或EAR01433,或由基因库登录号:NZ_AAOC01000003编码)的LLO蛋白。可以使用的LLO蛋白可以包括以下、基本上由以下组成或由以下组成:上述LLO蛋白中的任何LLO蛋白,或上述LLO蛋白的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段和同工型的片段。
也可以使用与LLO同源的蛋白质或其同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段以及同工型的片段。一个此类实例是蜂房杆菌溶素(alveolysin),所述蜂房杆菌溶素可以在例如蜂房芽孢杆菌(参见例如,基因库登录号:P23564或AAA22224,或由基因库登录号:M62709编码)中找到。其它此类同源蛋白是已知的。
LLO肽可以是全长LLO蛋白或截短的LLO蛋白或LLO片段。同样,LLO肽可以是保留天然LLO蛋白的一种或多种功能或缺乏天然LLO蛋白的一种或多种功能的肽。例如,保留的LLO功能可以允许细菌(例如,李斯特菌)从吞噬体或吞噬溶酶体中逃逸,或者增强与其融合的肽的免疫原性。保留的功能还可以是溶血性功能或抗原功能。可替代地,LLO肽可以是非溶血性LLO。LLO的其它功能以及用于评估LLO功能的方法和测定都是已知的。
LLO片段可以是PEST样序列或可以包括PEST样序列。LLO片段可以包括内部缺失、从C端截短和从N端截短中的一项或多项。在一些情况下,LLO片段可以包括一个以上的内部缺失。其它LLO肽可以是具有一种或多种突变的全长LLO蛋白。
一些LLO蛋白或片段相对于野生型LLO具有降低的溶血活性,或者是非溶血性片段。例如,可以通过羧基端的活化结构域的缺失或突变、通过半胱氨酸484的缺失或突变或通过另一位置处的缺失或突变来使LLO蛋白变为非溶血性的。
如US 8,771,702中详述的,通过胆固醇结合结构域(CBD)的缺失或突变使其它LLO蛋白变为非溶血性的,所述文献出于所有目的通过引用整体并入本文。突变可以包括例如取代或缺失。整个CBD可以进行突变,CBD的部分可以进行突变,或者CBD内的特定残基可以进行突变。例如,当与SEQ ID NO:55进行最佳比对时,LLO蛋白可以包括SEQ ID NO:55的残基C484、W491和W492(例如,C484、W491、W492、C484和W491、C484和W492、W491和W492或所有三个残基)以及对应残基中的一个或多个残基的突变(例如,对应的半胱氨酸或色氨酸残基)。作为实例,可以产生突变LLO蛋白,其中LLO的残基C484、W491和W492被丙氨酸残基取代,相对于野生型LLO,这将大大降低溶血活性。具有C484A、W491A和W492A突变的突变LLO蛋白被称为“mutLLO”。
作为另一个实例,可以产生具有包括胆固醇结合结构域的内部缺失的突变LLO蛋白。SEQ ID NO:55的胆固醇结合结构域的序列在SEQ ID NO:74中示出。例如,内部缺失可以是1-11个氨基酸缺失、11-50个氨基酸缺失或更长的氨基酸缺失。同样,突变的区域可以是1-11个氨基酸、11-50个氨基酸或更长(例如,1-50个、1-11个、2-11个、3-11个、4-11个、5-11个、6-11个、7-11个、8-11个、9-11个、10-11个、1-2个、1-3个、1-4个、1-5个、1-6个、1-7个、1-8个、1-9个、1-10个、2-3个、2-4个、2-5个、2-6个、2-7个、2-8个、2-9个、2-10个、3-4个、3-5个、3-6个、3-7个、3-8个、3-9个、3-10个、12-50个、11-15个、11-20个、11-25个、11-30个、11-35个、11-40个、11-50个、11-60个、11-70个、11-80个、11-90个、11-100个、11-150个、15-20个、15-25个、15-30个、15-35个、15-40个、15-50个、15-60个、15-70个、15-80个、15-90个、15-100个、15-150个、20-25个、20-30个、20-35个、20-40个、20-50个、20-60个、20-70个、20-80个、20-90个、20-100个、20-150个、30-35个、30-40个、30-60个、30-70个、30-80个、30-90个、30-100个或30-150个氨基酸)。例如,由SEQ ID NO:55的残基470-500、470-510或480-500组成的突变的区域将产生包括CBD(SEQ ID NO:55的残基483-493)的缺失序列。然而,突变的区域也可以是CBD的片段或可以与CBD的一部分重叠。例如,突变的区域可以由SEQ IDNO:55的残基470-490、480-488、485-490、486-488、490-500或486-510组成。例如,可以用异源序列替换CBD的片段(残基484-492),相对于野生型LLO,这将大大降低溶血活性。例如,可以用来自抗原NY-ESO-1(ESLLMWITQCR;SEQ ID NO:75)的CTL表位替换CBD(ECTGLAWEWWR;SEQ ID NO:74),所述CTL表位含有来自NY-ESO-1的HLA-A2限制性表位157-165。所得的LLO被称为“ctLLO”。
在一些突变的LLO蛋白中,突变的区域可以被异源序列替换。例如,突变的区域可以被相等数量的异源氨基酸、较小数量的异源氨基酸或较大数量的氨基酸(例如,1-50个、1-11个、2-11个、3-11个、4-11个、5-11个、6-11个、7-11个、8-11个、9-11个、10-11个、1-2个、1-3个、1-4个、1-5个、1-6个、1-7个、1-8个、1-9个、1-10个、2-3个、2-4个、2-5个、2-6个、2-7个、2-8个、2-9个、2-10个、3-4个、3-5个、3-6个、3-7个、3-8个、3-9个、3-10个、12-50个、11-15个、11-20个、11-25个、11-30个、11-35个、11-40个、11-50个、11-60个、11-70个、11-80个、11-90个、11-100个、11-150个、15-20个、15-25个、15-30个、15-35个、15-40个、15-50个、15-60个、15-70个、15-80个、15-90个、15-100个、15-150个、20-25个、20-30个、20-35个、20-40个、20-50个、20-60个、20-70个、20-80个、20-90个、20-100个、20-150个、30-35个、30-40个、30-60个、30-70个、30-80个、30-90个、30-100个或30-150个氨基酸)替换。其它突变的LLO蛋白具有一个或多个点突变(例如,1个残基、2个残基、3个残基或更多个残基的点突变)。突变的残基可以是连续的或不连续的。
在一个示例实施例中,LLO肽可以在信号序列中具有缺失,并且在CBD中具有突变或取代。
一些LLO肽是N端LLO片段(即,具有C端缺失的LLO蛋白)。一些LLO肽的长度为至少494个、489个、492个、493个、500个、505个、510个、515个、520个或525个氨基酸或长度为492-528个氨基酸。例如,LLO片段可以由LLO蛋白的大约前440个或441个氨基酸组成(例如,SEQ ID NO:55或56的前441个氨基酸,或者当与SEQ ID NO:55或56进行最佳比对时,另一个LLO蛋白的对应片段)。其它N端LLO片段可以由LLO蛋白的前420个氨基酸组成(例如,SEQ IDNO:55或56的前420个氨基酸,或者当与SEQ ID NO:55或56进行最佳比对时,另一个LLO蛋白的对应片段)。其它N端片段可以由LLO蛋白的约氨基酸20-442组成(例如,SEQ ID NO:55或56的氨基酸20-442,或者当与SEQ ID NO:55或56进行最佳比对时,另一个LLO蛋白的对应片段)。其它N端LLO片段包括不具有包括半胱氨酸484的活化结构域(并且具体地说,不具有半胱氨酸484)的任何ΔLLO。例如,N端LLO片段可以对应于LLO蛋白的前425个、400个、375个、350个、325个、300个、275个、250个、225个、200个、175个、150个、125个、100个、75个、50个或25个氨基酸(例如,SEQ ID NO:55或56的前425个、400个、375个、350个、325个、300个、275个、250个、225个、200个、175个、150个、125个、100个、75个、50个或25个氨基酸,或者当与SEQ ID NO:55或56进行最佳比对时,另一个LLO蛋白的对应片段)。优选地,片段包括一个或多个PEST样序列。LLO片段和截短的LLO蛋白可以含有对应于上述特定氨基酸范围中任何一个范围的同源LLO蛋白的残基。残基数量不必与上文列举的残基数量完全一致(例如,如果同源LLO蛋白相对于本文所公开的特定LLO蛋白具有插入或缺失)。N端LLO片段的实例包含SEQ ID NO:57、58和59。可以使用的LLO蛋白包括以下、基本上由以下组成或由以下组成:SEQ ID NO:57、58和59中所示的序列,或SEQ ID NO:57、58和59的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段和同工型的片段。在一些组合物和方法中,使用SEQ ID NO:59中所示的N端LLO片段。对SEQ ID NO:59中所示的N端LLO片段进行编码的核酸的实例是SEQ ID NO:60。
(2)ActA
可以在本文所公开的组合物和方法中利用的含PEST的肽的另一个实例是ActA肽。ActA是一种表面相关蛋白,并且在感染的宿主细胞中充当支架,以促进宿主肌动蛋白聚合物的聚合、组装和活化,从而通过细胞质推动单核细胞增生性李斯特菌。单核细胞增生性李斯特菌进入哺乳动物细胞的细胞溶质后不久,就会诱导宿主肌动蛋白丝的聚合,并且使用肌动蛋白聚合产生的力先在细胞内移动,然后在细胞之间移动。ActA负责介导肌动蛋白成核和基于肌动蛋白的动力。ActA蛋白为宿主细胞骨架组分提供了多个结合位点,从而充当了用于组装细胞肌动蛋白聚合机制的支架。ActA的N端与单体肌动蛋白结合,并且通过刺激内在肌动蛋白成核活性而充当组成型活性成核促进因子。actA基因和hly基因两者都是由转录活化因子PrfA调节的10-kb基因簇的成员,并且actA在哺乳动物细胞溶质中被上调大约226倍。可以使用对ActA蛋白或ActA蛋白的同源物、变体、同工型、类似物、同源物的片段、变体的片段或类似物的片段进行编码的任何序列。同源ActA蛋白与参考ActA蛋白的序列同一性可以为例如大于70%、72%、75%、78%、80%、82%、83%、85%、87%、88%、90%、92%、93%、95%、96%、97%、98%或99%。
ActA蛋白的一个实例包括SEQ ID NO:61中所示的序列、基本上由所述序列组成或由所述序列组成。ActA蛋白的另一个实例包括SEQ ID NO:62中所示的序列、基本上由所述序列组成或由所述序列组成。与这些序列中的任何一个序列相对应的前蛋白的前29个氨基酸是信号序列,并且当由细菌分泌时,其会与ActA蛋白切割开。ActA肽可以包括信号序列(例如,SEQ ID NO:61或62的氨基酸1-29),或者可以包括不包含信号序列的肽。ActA蛋白的其它实例包括以下、基本上由以下组成或由以下组成:SEQ ID NO:61或62的同源物、变体、同工型、类似物、片段、同源物的片段、同工型的片段或类似物的片段。
ActA蛋白的另一个实例是来自以下各项的ActA蛋白:单核细胞增生性李斯特菌10403S菌株(基因库登录号:DQ054585)、NICPBP 54002菌株(基因库登录号:EU394959)、S3菌株(基因库登录号:EU394960)、NCTC 5348菌株(基因库登录号:EU394961)、NICPBP 54006菌株(基因库登录号:EU394962)、M7菌株(基因库登录号:EU394963)、S19菌株(基因库登录号:EU394964)或单核细胞增生性李斯特菌的任何其它菌株。可以使用的LLO蛋白可以包括以下、基本上由以下组成或由以下组成:上述LLO蛋白中的任何LLO蛋白,或上述LLO蛋白的同源物、变体、同工型、类似物、片段、同源物的片段、变体的片段、类似物的片段和同工型的片段。
ActA肽可以是全长ActA蛋白或截短的ActA蛋白或ActA片段(例如,去除了C端部分的N端ActA片段)。优选地,截短的ActA蛋白包括至少一个PEST序列(例如,一个以上PEST序列)。另外,截短的ActA蛋白可以任选地包括ActA信号肽。截短的ActA蛋白中含有的PEST样序列的实例包含SEQ ID NO:45-48。一些此类截短的ActA蛋白包括SEQ ID NO:45-48中所示的PEST样序列中的至少两个PEST样序列或其同源物、SEQ ID NO:45-48中所示的PEST样序列中的至少三个PEST样序列或其同源物或SEQ ID NO:45-48中所示的PEST样序列中的所有四个PEST样序列或其同源物。截短的ActA蛋白的实例包含这样的蛋白质:其包括全长ActA蛋白序列(例如,SEQ ID NO:62)的约残基30-122、约残基30-229、约残基30-332、约残基30-200或约残基30-399,基本上由所述残基组成或由所述残基组成。截短的ActA蛋白的其它实例包含这样的蛋白质:其包括全长ActA蛋白序列(例如,SEQ ID NO:62)的大约前50个、100个、150个、200个、233个、250个、300个、390个、400个或418个残基,基本上由所述残基组成或由所述残基组成。截短的ActA蛋白的其它实例包含这样的蛋白质:其包括全长ActA蛋白序列(例如,SEQ ID NO:62)的约残基200-300或残基300-400,基本上由所述残基组成或由所述残基组成。例如,截短的ActA由如US 7,655,238中所描述的野生型ActA蛋白的前390个氨基酸组成,所述文献出于所有目的通过引用整体并入本文。作为另一个实例,截短的ActA可以是ActA-N100或其经修饰的版本(被称为ActA-N100*),其中PEST基序已经缺失并且含有如US 2014/0186387中所描述的非保守性QDNKR(SEQ ID NO:73)取代,所述文献出于所有目的通过引用整体并入本文。可替代地,截短的ActA蛋白可以含有与上文的氨基酸范围或本文所公开的ActA肽中的任何ActA肽的氨基酸范围中的一个范围相对应的同源ActA蛋白的残基。残基数量不必与本文列举的残基数量完全一致(例如,如果同源ActA蛋白相对于本文所利用的ActA蛋白具有插入或缺失,则残基数量可以相应地进行调节)。
截短的ActA蛋白的实例包含例如包括以下、基本上由以下组成或由以下组成的蛋白质:SEQ ID NO:63、64、65或66中所示的序列或SEQ ID NO:63、64、65和66的同源物、变体、同工型、类似物、变体的片段、同工型的片段或类似物的片段。SEQ ID NO:63被称为ActA/PEST1,并且由SEQ ID NO:62中所示的全长ActA序列的氨基酸30-122组成。SEQ ID NO:64被称为ActA/PEST2或LA229,并且由SEQ ID NO:62中所示的全长ActA序列中所示的全长ActA序列的氨基酸30-229组成。SEQ ID NO:65被称为ActA/PEST3,并且由SEQ ID NO:62中所示的全长ActA序列的氨基酸30-332组成。SEQ ID NO:66被称为ActA/PEST4,并且由SEQ IDNO:62中所示的全长ActA序列的氨基酸30-399组成。作为具体实例,可以使用由SEQ ID NO:64中所示的序列组成的截短的ActA蛋白。
截短的ActA蛋白的实例包含例如包括以下、基本上由以下组成或由以下组成的蛋白质:SEQ ID NO:67、69、70或72中所示的序列或SEQ ID NO:67、69、70和72的同源物、变体、同工型、类似物、变体的片段、同工型的片段或类似物的片段。作为具体实例,可以使用由SEQ ID NO:67中所示的序列(由SEQ ID NO:68中所示的核酸编码)组成的截短的ActA蛋白。作为另一个具体实例,可以使用由SEQ ID NO:70中所示的序列(由SEQ ID NO:71中所示的核酸编码)组成的截短的ActA蛋白。SEQ ID NO:71是单核细胞增生性李斯特菌10403S菌株中对ActA进行编码的前1170个核苷酸。在一些情况下,ActA片段可以与异源信号肽融合。例如,SEQ ID NO:72示出了与Hly信号肽融合的ActA片段。
C.产生对重组融合多肽进行编码的免疫疗法构建体
本文还提供了用于产生对包括本文所公开的重组融合多肽的组合物进行编码的免疫疗法构建体的方法。例如,此类方法可以包括:选择和设计抗原肽以包含在免疫疗法构建体中(以及例如,测试每个抗原肽的亲水性,并且如果抗原肽亲水性评分高于所选的亲水性指数阈值,则修饰或取消选择所述抗原肽);设计一种或多种包括所选的抗原肽中的每个抗原肽的融合多肽;以及产生对融合多肽进行编码的核酸构建体。
可以筛选抗原肽的疏水性或亲水性。可以选择抗原肽,例如,如果所述抗原肽是亲水性的,或者如果其评分达到或低于某个亲水性阈值,则这可以预测特定所关注的细菌(例如,单核细胞增生性李斯特菌)的分泌性。例如,可以通过具有21个氨基酸窗口的Kyte和Doolittle亲水性指数对抗原肽进行评分,排除所有高于临界值(约1.6)的评分,因为它们不太可能可由单核细胞增生性李斯特菌分泌。参见例如,Kyte-Doolittle(1982)《分子生物学杂志》157(1):105–132;所述文献出于所有目的通过引用整体并入本文。可替代地,可以更改评分约为所选临界值的抗原肽(例如,改变抗原肽的长度)。可以使用的其它滑动窗口大小包含例如9个、11个、13个、15个、17个、19个、21个、23个、25个、27个或更多个氨基酸。例如,滑动窗口大小可以是9-11个氨基酸、11-13个氨基酸、13-15个氨基酸、15-17个氨基酸、17-19个氨基酸、19-21个氨基酸、21-23个氨基酸、23-25个氨基酸或25-27个氨基酸。可以使用的其它临界值包含例如以下范围:1.2-1.4、1.4-1.6、1.6-1.8、1.8-2.0、2.0-2.2、2.2-2.5、2.5-3.0、3.0-3.5、3.5-4.0或4.0-4.5,或者临界值可以为1.4、1.5、1.7、1.8、1.9、2.0、2.1、2.2、2.3、2.3、2.5、2.6、2.7、2.8、2.9、3.0、3.1、3.2、3.3、3.4、3.5、3.6、3.7、3.8、3.9、4.0、4.1、4.2、4.3、4.4或4.5。临界值可以例如取决于用于递送融合多肽的细菌的属或种而变化。
其它合适的亲水性图或其它适当的量表包含例如在以下文献中进行报道的那些:Rose等人(1993)《生物物理和生物分子结构年鉴(Annu Rev Biomol Struct)》22:381-415;Biswas等人(2003)《色谱法期刊A(Journal of Chromatography A)》1000:637-655;Eisenberg(1984)《生物化学年鉴(Ann Rev Biochem)》53:595-623;Abraham和Leo(1987)《蛋白质:结构、功能和遗传学(Proteins:Structure,Function and Genetics)》2:130-152;Sweet和Eisenberg(1983)《分子生物学杂志》171:479-488;Bull和Breese(1974)《生物化学与生物物理学档案(Arch Biochem Biophys)》161:665-670;Guy(1985)《生物物理学杂志(Biophys J)》47:61-70;Miyazawa等人(1985)《高分子(Macromolecules)》18:534-552;Roseman(1988)《分子生物学杂志》200:513-522;Wolfenden等人(1981)《生物化学(Biochemistry)》20:849-855;Wilson(1981)《生物化学杂志(Biochem J)》199:31-41;Cowan和Whittaker(1990)《肽研究(Peptide Research)》3:75-80;Aboderin(1971)《国际生物化学与细胞生物学杂志(Int J Biochem)》2:537-544;Eisenberg等人(1984)《分子生物学杂志》179:125-142;Hopp和Woods(1981)《美国国家科学院院刊》78:3824-3828;Manavalan和Ponnuswamy(1978)《自然(Nature)》275:673-674;Black和Mould(1991)《分析生物化学(Anal Biochem)》193:72-82;Fauchere和Pliska(1983)《欧洲药物化学杂志(EurJ Med Chem)》18:369-375;Janin(1979)《自然》277:491-492;Rao和Argos(1986)《生物化学与生物物理学报(Biochim Biophys Acta)》869:197-214;Tanford(1962)《美国化学学会杂志(Am Chem Soc)》84:4240-4274;Welling等人(1985)《欧洲生化学会联合会快报》188:215-218;Parker等人(1986)《生物化学》25:5425-5431;以及Cowan和Whittaker(1990)《肽研究》3:75-80,所述文献中的每个文献出于所有目的通过引用整体并入本文。
任选地,可以对抗原肽与受试者人类白细胞抗原(HLA)类型结合的能力进行评分(例如,通过使用可在www.iedb.org获得的免疫表位数据库(IED),其包含netMHCpan、ANN、SMMPMBEC.SMM、CombLib_Sidney2008、PickPocket和netMHCcons),并且根据每个抗原肽的最佳MHC结合评分对其进行排名。其它来源包含TEpredict(tepredict.sourceforge.net/help.html)或其它可用的MHC结合测量量表。对于不同的表达载体(如沙门氏菌),临界值可能不同。
任选地,可以筛选抗原肽的免疫抑制表位(例如,T-reg表位、IL-10诱导性T辅助表位等),以取消选择抗原肽或避免免疫抑制的影响。
任选地,可以使用表位的免疫原性的预测算法来筛选抗原肽。然而,这些算法在预测哪种肽会产生T细胞应答时最多精确20%。可替代地,不使用筛选/预测算法。可替代地,可以筛选抗原肽的免疫原性。例如,这可以包括使一个或多个T细胞与抗原肽接触,并且分析免疫原性T细胞应答,其中免疫原性T细胞应答将肽鉴别为免疫原性肽。这还可以包括使用免疫原性测定法来测量CD25、CD44或CD69中的至少一种的分泌,或在使一个或多个T细胞与肽接触时,测量选自包括IFN-γ、TNF-α、IL-1和IL-2的组的细胞因子的分泌,其中增加的分泌将肽鉴别为包括一个或多个T细胞表位。
可以将所选的抗原肽排列成潜在融合多肽的一个或多个候选顺序。如果可用抗原肽多于单个质粒所能容纳的抗原肽,则可以根据需要/期望将不同的抗原肽分配优先级,和/或分成不同的融合多肽(例如,用于包含在不同的重组李斯特菌菌株中)。优先级可以由以下因素确定:如经翻译的多肽的相对大小、转录的优先级和/或总体疏水性。可以排列抗原肽,使得所述抗原肽在没有接头或者任何数量的抗原肽对之间的接头的任何组合的情况下直接连接在一起,如本文其它地方更详细地公开的。可以基于对需要的构建体的数量与突变负荷、来自单个质粒的多个表位的翻译和分泌效率以及包括质粒的每种细菌或Lm所需的MOI的考虑来确定要包含的线性抗原肽的数量。
可以对抗原肽或整个融合多肽(即,包括抗原肽和含PEST的肽以及任何标签)的组合的疏水性进行评分。例如,可以通过具有滑动的21个氨基酸窗口的Kyte和Doolittle亲水性指数对整个融合的抗原肽或整个融合多肽的亲水性进行评分。如果任何区域评分高于临界值(例如,约1.6),则可以在融合多肽内对抗原肽进行重新排序或改组,直到找到可接受的抗原肽顺序(即,其中没有区域评分高于临界值的抗原肽)。可替代地,可以去除任何有问题的抗原肽或将其重新设计为具有不同的大小。可替代地或另外地,可以添加或修饰本文其它各处所公开的抗原肽之间的一个或多个接头,以改变疏水性。与对单独的抗原肽的亲水性测试一样,可以使用其它窗口大小,或者可以使用其它临界值(例如,取决于用于递送融合多肽的细菌的属或种)。另外,可以使用其它适合的亲水性图或其它适当的量表。
任选地,可以进一步筛选抗原肽或整个融合多肽的组合的免疫抑制表位(例如,T-reg表位、IL-10诱导性T辅助表位等),以取消选择抗原肽或避免免疫抑制的影响。
然后可以设计和优化对抗原肽或融合多肽的候选组合进行编码的核酸。例如,可以针对增加的翻译水平、表达持续时间、分泌水平、转录水平以及其任何组合来优化序列。例如,相对于对照(非优化的序列),增加可以是2倍到1000倍、2倍到500倍、2倍到100倍、2倍到50倍、2倍到20倍、2倍到10倍或3倍到5倍。
例如,可以针对可能在寡核苷酸序列中形成的二级结构的降低的水平对融合多肽或编码融合多肽的核酸进行优化,或者可替代地,进行优化以防止可以修饰序列的任何酶的附着。细菌细胞中的表达可以例如通过转录沉默、低mRNA半衰期、二级结构形成、寡核苷酸结合分子(如阻遏物和抑制剂)的附着位点以及稀有的tRNA池的可用性而受到阻碍。在原始序列内发现了细菌表达中许多问题的来源。优化RNA可以包含修饰顺式作用元件、适应其GC含量、关于细菌细胞的非限制性tRNA池修饰密码子偏倚以及避免内部同源区域。因此,优化序列可能需要例如调节GC含量非常高(>80%)或者非常低(<30%)的区域。优化序列还可能需要例如避免以下顺式作用序列基序中的一个或多个:内部TATA盒、chi位点和核糖体进入位点;富含AT或富含GC的序列区段;重复序列和RNA二级结构;(隐蔽)剪接供体和受体位点;分支点;或其组合。优化表达还可能需要将序列元件添加到基因的侧翼区域和/或质粒中的其它地方。
优化序列还可能需要例如使密码子使用适应宿主基因(如单核细胞增生性李斯特菌基因)的密码子偏倚。例如,以下密码子可以用于单核细胞增生性李斯特菌。
表3.密码子。
| A=GCA | G=GGT | L=TTA | Q=CAA | V=GTT |
| C=TGT | H=CAT | M=ATG | R=CGT | W=TGG |
| D=GAT | I=ATT | N=AAC | S=TCT | Y=TAT |
| E=GAA | K=AAA | P=CCA | T=ACA | Stop=TAA |
| F=TTC |
可以产生对融合多肽进行编码的核酸,并且将其引入到递送媒剂(如细菌菌株或李斯特菌菌株)中。其它递送媒剂可以适用于DNA免疫疗法或肽免疫疗法,如牛痘病毒或病毒样颗粒。一旦产生对融合多肽进行编码的质粒并且将其引入到细菌菌株或李斯特菌菌株中,就可以培养细菌或李斯特菌菌株并且进行表征,以确认包括抗原肽的融合多肽的表达和分泌。
V.试剂盒
还提供了包括用于执行本文所公开的方法中的任何方法的一种或多种试剂的试剂盒,或包括本文所公开的任何组合物、工具或仪器的试剂盒。
例如,此类试剂盒可以包括THP-1细胞以及任选的一种或多种用于分化THP-1细胞的试剂或说明材料。此类试剂盒还可以包括本文所公开的重组细菌或李斯特菌菌株。另外,此类试剂盒可以另外包括说明材料,所述说明材料描述了THP-1细胞和/或重组细菌或李斯特菌菌株用于执行本文公开的方法的用途。尽管下文描述了模型试剂盒,但根据本公开,其它有用试剂盒的内容物是显而易见的。
出于所有目的,上文或下文引用的所有专利申请、网站、其它出版物、登录号等都通过引用整体并入,其程度如同每个单独的项目被单独并且具体地指出通过引用的方式并入。如果序列的不同版本与不同时间的登录号相关联,则意指在本申请的有效提交日期与该登录号相关联的版本。有效提交日期是指实际提交日期或提及登记号的优先权申请的提交日期(在适用情况下)中较早的日期。同样,如果出版物、网站等的不同版本在不同时间发布,除非另有说明,否则指在申请的有效提交日期最近发布的版本。除非另外具体说明,否则本发明的任何特征、步骤、元件、实施例或方面都可以与任何其它特征、步骤、元件、实施例或方面结合使用。尽管为了清楚和理解起见,已通过图解和实例方式详细地对本发明进行了描述,但显而易见的是,可以在所附权利要求的范围内进行某些改变和修改。
实施例清单
本文所公开的主题包含但不限于以下实施例。
1.一种评估测试李斯特菌菌株的减毒或感染性的方法,所述方法包括:
(a)用所述测试李斯特菌菌株感染分化的THP-1细胞,其中在用所述测试李斯特菌菌株感染之前,已将所述THP-1细胞分化为巨噬细胞;
(b)裂解所述THP-1细胞,并将所述裂解物平板接种在琼脂上;以及
(c)通过在所述琼脂上的生长来计数已在所述THP-1细胞内部繁殖的所述李斯特菌。
2.根据实施例1所述的方法,其进一步包括在步骤(a)之前,使用佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA)将所述THP-1细胞分化为巨噬细胞。
3.根据实施例1或2所述的方法,其中用所述测试李斯特菌菌株感染分化的THP-1细胞包括:用所述测试李斯特菌菌株接种所述分化的THP-1细胞,并将所述测试李斯特菌菌株与所述分化的THP-1细胞一起孵育1-5小时,以形成感染的THP1细胞。
4.根据前述实施例中任一项所述的方法,其中步骤(a)包括以1:1的感染复数(MOI)感染所述分化的THP-1细胞。
5.根据前述实施例中任一项所述的方法,其进一步包括在步骤(a)与(b)之间,杀死未被所述THP-1细胞摄取的李斯特菌。
6.根据实施例5所述的方法,其中使用抗生素进行所述杀死,任选地其中所述抗生素是庆大霉素。
7.根据实施例1到4中任一项所述的方法,其中在步骤(b)之前,从感染的THP-1细胞中去除细胞外李斯特菌。
8.根据实施例7所述的方法,其中去除细胞外李斯特菌包括添加对所述李斯特菌有效的抗生素,任选地其中所述抗生素是庆大霉素。
9.根据实施例7或8所述的方法,其中在去除细胞外李斯特菌之后和步骤(b)之前,将感染的THP-1细胞在生长培养基中孵育0-10小时。
10.根据前述实施例中任一项所述的方法,其中在感染后0小时进行步骤(b)。
11.根据前述实施例中任一项所述的方法,其中在感染后0小时、感染后1小时、感染后3小时和/或感染后5小时进行步骤(b)。
12.根据前述实施例中任一项所述的方法,其中所述琼脂含有能够支持所述李斯特菌生长的培养基。
13.根据前述实施例中任一项所述的方法,其进一步包括比较所述测试李斯特菌菌株与野生型李斯特菌菌株和/或参考样品的摄取和细胞内生长。
14.根据前述实施例中任一项所述的方法,其中所述测试李斯特菌菌株是单核细胞增生性李斯特菌菌株。
15.根据前述实施例中任一项所述的方法,其中所述测试李斯特菌菌株是重组李斯特菌菌株,所述重组李斯特菌菌株包括有包括对融合多肽进行编码的第一开放阅读框的核酸,其中所述融合多肽包括与疾病相关型抗原肽融合的含PEST的肽。
16.根据实施例15所述的方法,其中所述含PEST的肽是李斯特菌溶血素O(LLO)或其片段,并且所述疾病相关型抗原肽是人乳头瘤病毒(HPV)蛋白E7或其片段。
17.根据实施例15或16所述的方法,其中所述重组李斯特菌菌株是减毒的单核细胞增生性李斯特菌菌株,所述减毒的单核细胞增生性李斯特菌菌株包括prfA的缺失或prfA中的失活突变,其中所述核酸位于附加型质粒中并且包括对D133V PrfA突变蛋白进行编码的第二开放阅读框。
18.根据实施例15所述的方法,其中所述重组李斯特菌菌株是减毒的单核细胞增生性李斯特菌菌株,所述减毒的单核细胞增生性李斯特菌菌株包括actA、dal和dat的缺失或actA、dal和dat中的失活突变,其中所述核酸位于附加型质粒中并且包括对丙氨酸消旋酶或D-氨基酸氨基转移酶进行编码的第二开放阅读框,并且其中所述含PEST的肽是李斯特菌溶血素O(LLO)的N端片段。
19.一种评估测试细菌菌株的减毒或感染性的方法,所述方法包括:
(a)分化THP-1细胞;
(b)用所述测试细菌菌株感染所述分化的THP-1细胞,其中所述感染包括:
(i)用所述测试细菌菌株接种所述分化的THP-1细胞;
(ii)将所述测试细菌菌株与所述分化的THP-1细胞一起孵育1-5小时,以形成感染的THP1细胞;
(iii)从所述感染的THP-1细胞中去除细胞外细菌;以及
(iv)将所述感染的THP-1细胞在生长培养基中孵育0-10小时;
(c)裂解所述感染的THP-1细胞以形成裂解物;
(d)将所述裂解物或所述裂解物的稀释液平板接种在含有能够支持所述细菌生长的培养基的板上;以及
(e)计数所述板上的所述细菌的菌落形成单位。
20.根据实施例19所述的方法,其中感染所述分化的THP-1细胞的步骤是以1:1的感染复数(MOI)进行的。
21.根据实施例19或20所述的方法,其中去除细胞外细菌的步骤包括添加对所述细菌有效的抗生素,任选地其中所述抗生素是庆大霉素。
22.根据实施例19到21中任一项所述的方法,其中将所述感染的THP-1细胞在生长培养基中孵育0小时、1小时、3小时或5小时。
23.根据实施例19到22中任一项所述的方法,其中所述测试细菌菌株是单核细胞增生性李斯特菌菌株。
序列简要说明
使用核苷酸碱基的标准字母缩写和氨基酸的三字母代码示出随附序列表中列出的核苷酸和氨基酸序列。核苷酸序列遵循从序列的5'端开始并且向前(即,在每行中从左到右)到达3'端的标准惯例。每个核苷酸序列仅示出一条链,但任何提及的显示链均应理解为包含互补链。氨基酸序列遵循从序列的氨基端开始并且向前(即,在每行中从左到右)到达羧基端的标准惯例。
表4.序列说明。



实例
实例1.用于对单核细胞增生性李斯特菌的细胞内生长进行定量的基于THP-1的测定。
此实施例提供了用于定量野生型单核细胞增生性李斯特菌和减毒的重组单核细胞增生性李斯特菌的感染速率和/或细胞内生长的方法。使用分化的THP-1细胞进行的基于细胞的测定法用于分析基于李斯特菌的免疫疗法的细胞内生长,通过在脑心浸液琼脂上的生长来定量感染后的细菌。在一些实施例中,所描述的程序适用于ADXS11-001或其它李斯特菌菌株的样品。
单核细胞增生性李斯特菌是造成人类李斯特菌病的革兰氏阳性、无芽孢形成的细菌生物体。单核细胞增生性李斯特菌通过从人类巨噬细胞内的吞噬体逃逸而在体内存活。一旦逃逸,单核细胞增生性李斯特菌就能够在其宿主的细胞溶质内进行细胞内复制。免疫疗法菌株Lm-LLOE7(例如,ADXS11-001单核细胞增生性李斯特菌,一种活的减毒菌株)含有用于表达所关注的重组蛋白(即,与截短的李斯特菌溶血素O(tLLO)融合的人乳头瘤病毒蛋白E7)的质粒。Lm-LLOE7免疫疗法中使用的细菌菌株是突变菌株XFL-7,其缺少必需的毒力基因prfA。prfA基因是作用于包含所有毒力基因(如actA和hly,对LLO进行编码的基因)的多个基因的转录因子,但其对于李斯特菌的体外培养是不需要的。XFL-7是无毒的,并且可以被巨噬细胞摄取,但不能逃逸吞噬体以在巨噬细胞的细胞溶质中繁殖。为了评估Lm-LLOE7的减毒,在巨噬细胞感染测定中与野生型单核细胞增生性李斯特菌同时评估感染和复制。
重组蛋白由质粒pGG55表达,所述质粒含有在hly启动子控制下的无活性LLO和HPVE7编码序列的融合体,所述启动子还驱动prfA的质粒拷贝的表达。这些基因被引入到革兰氏阳性/革兰氏阴性细菌穿梭质粒pAM401中,所述质粒可以在大肠杆菌和李斯特菌中扩增,因为在革兰氏阳性生物体中不易进行基因操纵。因此,质粒基因包含革兰氏阳性和革兰氏阴性细菌的复制因子,以及革兰氏阳性和革兰氏阴性细菌的抗生素选择标志物(氯霉素)。质粒赋予了对氯霉素的抗性,并且通过在氯霉素存在下的培养进行体外保持。在体内,通过在XFL-7中失活的毒力因子PrfA的反式互补来保留质粒。
本文描述了使用分化的THP-1细胞来分析基于李斯特菌的免疫疗法的细胞内生长的基于细胞的测定法。THP-1细胞是可以通过用佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA)刺激而分化为巨噬细胞的人单核细胞。通过裂解THP-1细胞并且将细菌稀释液平板接种在脑心浸液琼脂上,在具体时间点对细菌进行感染前和感染后定量。菌落形成单位(CFU)表示在巨噬细胞细胞内环境中存活的活生物体。
下文阐述了使用ADXS11-0001的示例性程序。然而,这些程序和在其它实例中描述的程序可以用于任何李斯特菌菌株。感染前,将一个或多个样品和参考标准品解冻、沉淀、重悬并且稀释至目标细胞数。
表5.示例性材料。

表6.示例性设备(Apparatus/Equipment)。

化学品/试剂
在此测定中使用之前,可以目测检查BHI板,以确保没有大的污染以及琼脂的均匀扩散。通过用野生型10403S和ADXS11-001划线并在37℃下孵育24小时,可以检查板的生长适合性。野生型和ADXS11-001两者的菌落均应可见。
表7.试剂。
| RPMI 1640(西格玛(Sigma),目录号R8758或等效物) |
| FBS(西格玛,目录号F0926或等效物) |
| L-谷氨酰胺200mM(Cellgro,目录号25-005-CL或等效物) |
| 佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA),西格玛,目录号P8139或等效物 |
| DMSO(Amresco,目录号67-68-5或等效物) |
| 10mg/mL庆大霉素(西格玛,目录号221465或等效物) |
| 25mg/mL氯霉素(Amresco(VWR),目录号56-75-7或等效物) |
| 脑心浸液琼脂板(BD,目录号PA-255003.08或等效物) |
| PBS-不含钙和镁(Fisher,目录号10010-023或等效物) |
| 无菌水(WFI)(Fisher,目录号BP2470-1或等效物) |
| 野生型:单核细胞增生性李斯特菌(Lm)(PHE培养物收集) |
| THP1细胞系:西格玛,目录号88081201 |
| 链霉素,100mg/mL(西格玛奥德里奇(Sigma-Aldrich)56501或等效物) |
| 注射用无菌水,目录号BP281-1或等效物 |
| 当前ADXS11-001参考标准品 |
试剂制备。可以调整所有试剂调配物,以满足需要的所需体积。
完整的RPMI(c-RPMI)
1.向445mL的RPMI中添加以下内容:(1)50mL的FBS-辐照的;(2)5mL的L-谷氨酰胺(200mM)。
2.标记并储存在5±3℃下。有效期为自制备日期起1个月。
1.6mM PMA(佛波醇12-肉豆蔻酸酯13-乙酸酯)
1.在1mL的DMSO中重新配制1mg的PMA,使最终浓度为1.6mM PMA。
2.将10μL等分分装到微量离心管中直至耗尽。
3.标记并储存在-20±10℃下。有效期为自制备日期起6个月。
25μg/mL氯霉素
1.在20mL的100%乙醇中重新配制0.5g的氯霉素,使最终浓度为25μg/mL氯霉素。
2.标记并储存在-20±10℃下。有效期为自制备日期起1个月。
100μg/mL链霉素
1.在40mL的无菌水中重新配制4g的链霉素,使最终浓度为100μg/mL链霉素。使用0.2微米过滤器灭菌。将1mL等分分装到1.5mL试管中直至耗尽。
2.标记并储存在-20±10℃下。有效期为自制备日期起1个月。
脑心浸液琼脂+25μg/mL氯霉素
1.在进行之前,验证BHI板没有制造缺陷(污染、板破裂、琼脂不均匀等)。
2.将180μL的无菌PBS和20μL的氯霉素(25mg/mL)添加到脑心浸液琼脂板,并且使用无菌涂布器涂布以覆盖板的整个表面。涂布直到琼脂板吸收所有液体。
3.如果制备不止一个板,则可以制备氯霉素的工作储备液(180μL×PBS的板数和20μL×氯霉素的板数),并且向每个板中添加200μL,并且使用无菌涂布器涂布。
4.BHI+25μg/mL氯霉素板的有效日期将是按照制造商的板有效日期或氯霉素储备液的有效日期,以最早者为准。
脑心浸液琼脂+100μg/mL链霉素
1.在进行之前,验证BHI板没有制造缺陷(污染、板破裂、琼脂不均匀等)。
2.将180μL的无菌PBS和20μL的链霉素(100mg/mL)添加到脑心浸液琼脂板,并且使用无菌涂布器涂布以覆盖板的整个表面。涂布直到琼脂板吸收所有液体。
3.如果制备不止一个板,则可以制备链霉素的工作储备液(180μL×PBS的板数和20μL×链霉素的板数),并且向每个板中添加200μL,并且使用无菌涂布器涂布。
4.BHI+100μg/mL链霉素板的有效日期将是按照制造商的板有效日期或链霉素储备液的有效日期,以最早者为准。
对照
阴性/无菌对照
1.未接种的:每个三块板,BHI琼脂+链霉素100μg/mL和BHI琼脂+25μg/mL氯霉素。
2.接种的:每个三块板,各自用100μL的PBS接种,BHI琼脂+链霉素100μg/mL和BHI琼脂+25μg/mL氯霉素
阳性对照
1.野生型:在BHI琼脂+链霉素100μg/mL上划线有单核细胞增生性李斯特菌(PHE培养物收集10403S)的两个板。
2.ADXS11-001:用ADXS11-001参考标准品划线的两个板将作为BHI琼脂+25μg/mL氯霉素的阳性对照
THP-1细胞的制备
1.按照程序步骤3的要求解冻足够的THP-1细胞小瓶。
2.解冻后至少两次更替继代接种。在一些实施例中,THP-1细胞的传代次数低于32。
3.已经培养的THP-1细胞可以与适当的细胞培养参考物一起用于测定。
4.在c-RPMI中以1.0×106个细胞/mL的浓度制备40mL细胞。确定细胞计数。
5.向24孔板上的两个孔中的每个孔中添加1mL的细胞悬液(对于一个实例,参见附录2表9中的板图)。将孔标记为“无PMA”。向剩余的细胞悬液(大约34mL)中添加16μM PMA(34μL),使最终浓度为大约16nM PMA。均匀混合。
6.每孔分配1mL,总共10孔(例如,参见附录2表9中的板图)。
7.在36±1℃、5±1%CO2下孵育过夜(16-20小时)。
感染。对阳性对照野生型细菌执行以下步骤1-13,并且对于参考标准品和样品细菌将重复以下步骤。
1.适当地取出一小瓶阳性对照野生型单核细胞增生性李斯特菌或参考标准品或样品单核细胞增生性李斯特菌。
2.为了完全解冻小瓶,在36±2℃下孵育1分钟,然后在室温下孵育5分钟。
3.用注射器和针头将总体积转移至分别标记的1.5mL离心管。
4.将1.0mL转移至分别标记的1.5mL离心管。丢弃残留材料。
5.使用微量离心机以16,100×g沉淀1.0mL的细胞,持续2分钟。丢弃上清液,并用1.0mL的室温c-RPMI重悬细胞。使用c-RPMI制备细菌稀释液,得到最终浓度为1.0×106CFU/mL。此浓度下的最终体积应为大约15mL。
6.获得含有THP-1细胞的24孔板。适当地将孔标记为野生型或带有样品编号。
7.在显微镜下观察细胞,并确认用PMA处理的细胞与未经处理的细胞之间的区别。未经处理的细胞在轻轻摇动时会表现出流动性。轻轻摇动后,经处理的细胞将保持粘附。
8.使用移液管从所有含有经PMA处理的细胞的孔中吸出培养基(可以使用真空辅助)。
9.将1.0mL制备好的细菌(步骤6;1×106CFU/mL)转移到板的孔中。
10.在显微镜上观察板以确保THP-1细胞仍粘附在孔表面。
11.在36±2℃、5±1%CO2下孵育板。记录孵育开始时间。将板孵育2小时,然后再进行下一步操纵。
12.对1×106CFU/mL细菌稀释液(指定为p-2)进行生存力测试。利用附录1中概述的程序。利用附录1中概述的稀释方案。制备附录1中概述的阳性对照和阴性对照。
13.使用测试单核细胞增生性李斯特菌样品(例如,ADXS11-001)重复步骤1-12。
感染停止。将首先对阳性对照野生型细菌执行以下步骤1-10,并将对样品细菌重复以下步骤。
1.制备含有20μg/mL庆大霉素的c-RPMI。
2. 2小时后,从36±2℃、5±1%CO2孵育条件下去除含有野生型或样品的板。
3.使用移液管从每个孔中去除含有单核细胞增生性李斯特菌的培养基(可以使用真空辅助)。
4.每孔小心地分配1mL制备好的含有20μg/mL庆大霉素的c-RPMI,向孔的一侧缓慢添加以避免破坏。
5.将板放回孵育条件(36±2℃,5±1%CO2)中,持续42-45分钟。
6.从孵育条件中取出板。
7.使用移液管从每个孔中去除含有20μg/mL庆大霉素的c-RPMI(可以使用真空辅助)。
8.通过在每孔中添加1mL不含庆大霉素的c-RPMI(向孔的一侧缓慢添加以避免破坏),小心地洗涤细胞。
9.使用移液管从每个孔中去除c-RPMI(可以使用真空辅助)。
10.通过向孔的一侧缓慢添加以避免破坏而小心地将1mL的c-RPMI(不含庆大霉素)分配到每个孔中,并且将板放回36±2℃、5±1%CO2的孵育条件中,持续5-15分钟。
11.使用含有参考标准品的板重复步骤1-10,并使用含有样品(例如,ADXS11-001)的板重复。
用于检测细胞内单核细胞增生性李斯特菌生长的收集程序
1.从孵育条件中取出板,并记录时间。第一时间点将是p0。将采用的后续时间点为p3(3小时),以及任选的p5(5小时)。
2.在显微镜下观察孔。确认每个孔中经PMA处理的THP-1细胞的层是一致的,并且确认在之前的抽吸和分配步骤期间很少或没有细胞被移出和去除。如果观察到任何孔有显著的THP-1细胞丢失,则用“X”做好标记以提示不要使用这个孔。
3.选择一个孔用于时间点“p0”的收集。
4.通过用移液管抽吸,从所选的孔中去除c-RPMI(可以使用真空辅助)。
5.将1mL的无菌水分配到孔中,并且使用微量移液管,通过上下移液将THP-1细胞从孔表面移出。
6.将全部内容物转移到1.5mL离心管中。
7.在显微镜下观察孔以确认细胞已成功去除。如果仍然残留大量THP-1细胞,则利用先前转移到1.5mL管中的一部分无菌水通过上下移液移出细胞。将内容物转移回1.5mL管中,并通过显微镜术确认THP-1细胞已被去除。
8.将板放回孵育条件(36±2℃,5±1%CO2)中直到准备好收集下一个时间点为止。
9.将试管涡旋持续至少1分钟。
10.执行生存力测试。利用附录1中概述的程序。利用附录1表8中概述的稀释方案。
11.使用含有参考标准品的板重复步骤1-10,并且使用含有样品(例如,ADXS11-001)的板重复。
计算
1.摄取量(p-2/p0),表示为样品与野生型的比率。
2.细胞内生长(p3/p0),表示为野生型与样品的比率。
附录1-生存力测试程序
1.在启动生存力测试之前确保所有琼脂板都充分干燥。
2.制备以下阴性对照:(1)三个未接种的适当琼脂类型的板;以及(2)三个用100μL的PBS接种且用无菌涂布器涂布的适当琼脂类型的板。
3.将THP-1细胞的1.0mL等分试样以最大速度涡旋60秒。p-2时间点的生存力将改为利用在PBS中制备的1.0×106CFU/mL稀释液。
4.将基于单核细胞增生性李斯特菌细胞类型(野生型或样品)和所测试的时间点制备系列稀释液。参考表8。通过将100μL经涡旋的1.0mL等分试样转移到900μL的PBS中来制备系列稀释液。重复此过程,直到获得所需的所有稀释液为止。
5.将使用无菌涂布器将每种稀释液的接种物一式三份地涂布在适当的琼脂类型上。
6.制备以下阳性对照。用10μL接种环将适当的阳性对照一式两份接种到适当的琼脂上。
7.将使每个板吸收液体并盖上盖子干燥至少15分钟,然后将其倒置并且放置在35-38℃的孵育箱中。
8.16-24小时后,从孵育条件中取出板。确保在每个时间点将所有单核细胞增生性李斯特菌细胞类型(野生型和样品)孵育相同的持续时间。
9.将针对每种稀释液的每个板手动计数并记录菌落形成单位(CFU)的总数。
表8.待用于在每个时间点对野生型和样品进行生存力测试的稀释度(值可以根据需要进行调整)。

附录2-24孔板的制备。下文示出了24孔板的制备。对Lm野生型进行这种板设置,然后对参考标准品和样品(例如,ADXS11-001)重复进行此设置。可以基于接种时计数的THP-1细胞的数量和待测试的时间点来调整板孔设置。
表9. 24孔板设置。

实例2.用于对单核细胞增生性李斯特菌的细胞内生长进行定量的基于THP-1的测定法的验证。
进行此项资格研究以证明与野生型单核细胞增生性李斯特菌(Lm)相比,实例1中所描述的方法可以用于定量ADXS11-001药物产品的减毒。方法利用了人类THP-1细胞,并且评估了THP-1细胞中的ADXS11-001药物产品或野生型Lm的摄取和细胞内生长。此实例总结了从资格实验中产生的数据。
表10.总结-方法资格表。

单核细胞增生性李斯特菌是在抗原呈递细胞(APC)中表现出独特的生命周期的革兰氏阳性、无芽孢形成的细菌生物体。在APC吞噬体最初摄取Lm后,触发了溶细胞素、李斯特菌溶血素O(tLLO)的表达,这介导Lm从吞噬体中逃逸。一旦逃逸,Lm就能够在其宿主的细胞溶质内进行细胞内复制。使用分化的THP-1细胞进行的基于细胞的测定法用于分析基于李斯特菌的疫苗的摄取和细胞内生长。THP-1细胞是人类巨噬细胞,其在培养物中作为单核细胞保持,但可以通过用佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA)刺激而轻易分化为巨噬细胞。通过裂解THP-1细胞并且将细菌稀释液平板接种在脑心浸液琼脂上,在具体时间点对细菌进行感染前和感染后定量。菌落形成单位(CFU)表示在巨噬细胞细胞内环境中存活的活Lm。
菌株ADXS11-001含有用于表达所关注的蛋白质(即,与截短的李斯特菌溶血素O(tLLO)融合的人乳头瘤病毒蛋白E7)的质粒。THP-1感染测定法用于证明ADXS11-001相对于野生型亲本菌株10403S的减毒。在此测定中,以1:1的感染复数用10403S或ADXS11-001感染THP1细胞,并且在不同时间点(如感染后1小时、3小时和5小时)分析了细菌CFU的体外生长。减毒的结果是,与10403S相比,观察到ADXS11-001的摄取和细胞内生长显著降低。
对照的制备。如实例1所述制备野生型Lm 10403S和ADXS11-001 DP。简而言之,将样品在36±2℃下解冻并进行离心,并且使用完整的RPMI将浓度调节到1.0×106个细胞/mL。
THP-1细胞的制备。将THP-1细胞库(传代号P33)以1×106个活细胞/mL的密度冷冻。如实例1所述制备THP-1细胞;P33。简而言之,将THP-1细胞以1.0×106个细胞/mL/孔的浓度平板接种在含有16nM PMA的完整的RPMI中的24孔板中。
样品的制备。如实例1中那样,用野生型Lm 10403S和ADXS11-001 DP感染PMA分化的THP-1细胞。通过裂解THP-1细胞并且通过将细菌稀释液平板接种在琼脂板上,在具体时间点对细菌菌落形成单位(CFU)进行感染前和感染后定量。
结果。结果从三个独立的实验中产生。分析从每种稀释液产生的CFU以及对照和样品的每个时间点,以捕获所有所需的计算,并对资格参数进行评估。确定测定间精度和测定内精度的均值、标准差、变异系数和原始数据输出的计算,并评估每次运行的特异性。
以计数的细胞的数量表示的生存力充当此测定的原始数据输出。以下标准用于确定生存力。仅当阴性对照(未接种的板和经PBS接种的板)未示出菌落生长时,才将来自测定的数据视为可接受的。小于40的菌落形成单位(CFU)被视为计数太少(TFTC),而大于600的CFU被视为计数太多(TNTC)。仅对这些限制内的值进行定量。
精度(测定内重复性)
针对测定内精度,计算复制对照和样品的值的%相对标准偏差(RSD)。在每个时间点处,三个测定中的每个测定中的一式三份孔的%RSD的范围为:野生型,11%到20%;以及ADXS11-001参考标准样品,9%到21%。对于野生型和ADXS11-001参考标准品两者,跨所有时间点通过%RSD测得的最大测定内变异为21%,并且所述变异其在p1时间点观察到。然而,p1值未用于计算可报告的结果。预计测定内精度正好处于21%RSD之内。
用于计算测定的可报告值的p0的值为:野生型,20%RSD最大值;以及参考标准品,9%RSD最大值。细胞内生长输出p3和p5分别显示:野生型的最大%RSD为11%和17%,而ADXS11-001参考标准品的最大%RSD为10%和19%。p3值示出的测定间精度高于p5值。表11总结了测定内精度的RSD值。另外,如图1中的曲线所示,以时间对活细胞计数(VCC)作图的生长速率在孔之间没有示出明显差异,因为所有曲线均示出相同的总体形状和趋势。
表11.在资格测定中的每个资格测定的每个时间点处的%RSD值。

中间精度。
使用由两名分析员在多天内进行的三个独立测定获得的值来评估中间精度。进行了利用三个THP-1细胞传代次数以及感染和滴定的三个测定。评估了以变异系数表示的单独的测试结果之间的一致性程度,包含根据每个独立测定制备在每个感染后时间点处,样品的三个复制测量结果的平均值之间的一致性。评估还包含根据每个独立测定制备在每个感染后时间点处,野生型对照的三个复制测量结果的平均值之间的一致性。
(A)原始数据,每个时间点处的VCC。对于所有三个种测定,均计算了在每个时间点针对稀释度归一化的VCC值。观察到野生型的最高%RSD为47%,而ADXS11-001参考标准品的最高%RSD为23%。结果总结在表12中。应当注意的是,原始数据的%RSD并不那么重要,因为对于可报告的结果,这些值进一步被转换为比率。
另外,如图2中的曲线所示,以时间对活细胞计数(VCC)作图的生长速率在测定之间没有示出明显差异,因为所有曲线均示出相同的总体形状和趋势。
表12.针对资格测定中的每个资格测定的每个时间点的稀释度归一化的原始数据VCC。

(B)测定比率(可报告值)。对于每个实验,可报告值如下进行计算:
样品摄取:
细胞内生长:
细胞内生长:
对于所有三个测定,样品摄取的比率大于10,而细胞内生长的比率大于2。在所有三个测定之间,样品摄取的最大%RSD为39%,而细胞内生长的最大%RSD为29%,所述最大%RSD是可报告的比率值。结果示于表13中。另外,如图1中的曲线所示,以时间对活细胞计数(VCC)作图的生长速率在测定之间没有示出明显差异,因为所有曲线均示出相同的总体形状和趋势。
表13.来自三个资格测定的可报告值的结果。

(C)分析员。对于测定3和4的测定的感染部分由测定5中的分析员1和分析员2进行。对于测定3和4的测定的滴定部分由测定5中的分析员2和分析员1进行。数据表明分析员之间的野生型原始数据值可能存在差异,并且这反映在p-2/p0比率中。可报告值p3/p0和p5/p0没有示出分析员的影响。独立于分析员的测定比率对于摄取而言为12或更高而对于细胞内生长而言大于3,这是足以区分野生型菌株与ADXS11-001参考标准品或样品的倍数差异。参见表13。
(D)日期。对于在不同日期进行的测定的可报告值,未观察到显著影响。测定3和测定4(其中测定的滴定和感染部分由同一分析员进行)的可报告值对于摄取而言在3个单位(p-2/p0)而对于细胞内生长而言在2个单位之内。所有值对于摄取而言为12或更高而对于细胞内生长而言大于2,这是明显足以区分野生型菌株与ADXS11-001参考标准品和样品的倍数差异。参见表13。
(E)THP-1细胞的传代次数。细胞传代次数对测定的可报告值没有显著影响。THP1细胞传代P32、P37和P39用于此项资格。每个传代次数给出的可报告值为对于摄取而言为12或更高而对于细胞内生长而言大于2,这是明显足以区分野生型菌株与ADXS11-001参考标准品和样品的倍数差异。参见表13。
特异性
测试了未接种的和经PBS接种的THP-1基质空白样品的干扰和选择性。这包含在每个测定中,并且在所有测定中均未观察到空白的生长(无CFU)。这些阴性对照还证明不存在污染,这表明没有假阴性或假阳性结果。
在所述测定中的每个测定中,在样品和野生型对照两者中均产生了来自裂解的THP-1基质的可检测CFU。在参考标准样品(ADXS11-001)和对照(野生型)的相同的THP-1基质的存在下,从所述测定中的每个测定中均检测到CFU,从而证明了可接受的特异性。
另外,细胞内生长是重组Lm疫苗株能够进入细胞并且因此繁殖从而支持选择性的指标。针对三个测定中的每个测定,观察并计算了细胞内生长,这足以证明野生型菌株与ADXS11-001参考标准品和样品之间的倍数差异。
结果。实例1中所示出的方案已被鉴定为有资格用于在ADXS11-001的分化的THP-1细胞中对单核细胞增生性李斯特菌感染和复制进行分析。方法被证明是特异性的,因为所述方法检测到野生型与ADXS11-001在摄取和细胞内生长方面的倍数差异。所述方法还被表明是精确且可重复的,并且可报告的测定结果类似地独立于分析员、进行测定的日期或THP-1细胞的传代次数。
实例3.用于对单核细胞增生性李斯特菌的细胞内生长进行定量的基于THP-1的测定法的优化。
使用实例1示出的方法从总共13次代表性测试运行中获得数据。对数据进行了评估,以寻求方法效率的提高,同时保持关键的质量属性,包含确定用于发展应答的较短的时间框是否合理(3小时对5小时)、找到THP-1细胞的传代次数的上限、找到应答从p-2到p0的基线变化的下限以及确定p1时间点的效用。
主题方法是基于细胞的巨噬细胞感染测定法,其用于评估ADXS11-001的感染和复制,作为评估其减毒的一部分。并行使用野生型(WT)单核细胞增生性李斯特菌细胞10403S和特定ADXS11-001样品两者进行此测定。这是基于细胞的测定法,并且其使用的细菌可通过裂解THP-1细胞并且将细菌稀释液平板接种在琼脂上,在具体时间点进行感染前和感染后后定量。菌落形成单位(CFU)表示在巨噬细胞细胞内环境中存活的活生物体的计数。在不同时间点定量的CFU与自身和WT的比率为定量结果提供了机会。
作为方法的感染步骤的一部分,24孔板用于对样品和WT两者产生差异反应,然后将二者取样并且孵育,以获得活细胞计数。此活细胞计数被称为p-2,因为其在感染开始和2小时孵育时间之前,在此之后,再次对样品和WT进行测量。此2小时孵育后的测量结果被称为p0。在1小时(p1)、3小时(p3)和5小时(p5)孵育时间后,还进行了随后的活细胞计数测量。所述方法报告:(a)样品的更新(p-2/p0),表示为样品与WT的比率;以及(b)细胞内生长(p3/p0),表示为WT与样品的比率。数据来源在表14中示出。
表14.分析中使用的数据。

筛选符合预期的结果。图3显示了在本发明方法中的所有时间点(p-2、p0、p1、p3和p5)观察到的原始计数信息。表12中记录的运行中的每个运行都在单独的子图中,并且关键批次中的每个批次的所得曲线表示测试结果。数据证明了最初两个小时内(p-2到p0)的预期的下降变化,然后是从p0到p5的增加。
响应如预期一样,其中样品在初次接种后的计数明显低于野生型生物体,并且在p0时间点之后的增长速率类似。
图4和图5示出了相对于野生型(WT)的样品生长的摄取数据的图形描绘。图4示出了原始数据,表示为p-2处的计数与p0处看到的计数的比率。对于样品,从p-2到p0的变化量明显不同。图5示出了相同的内容,但是将样品结果转换为与野生型的比率。
图5显示,样品对比野生型的p-2/p0应答的变化的比率随运行而变化。相对于野生型,变化通常大于样品的5倍差异。这在图5中以红色虚线示出。样品对野生型的相对应答与THP-1细胞的传代次数有关(如下所示)。
图6和图7示出了在取得相对于野生型的比率之前的细胞内生长(p3/p0)和(p5/p0)的数据的图形描绘。图6显示了在取得相对于野生型的比率之前的样品比率。在取得比率之前,p3和p5处的结果存在显著差异。图7根据所述方法调整数据以示出相对于野生型的响应的变化。其显示p3、p5响应处的相对响应无显著差异。在样品类型内(两次运行之间)和样品之间观察到类似的可变性。
图8绘制了与图7所示相同的结果,但也按运行方式细分数据。此结果视图显示,在p3和p5处的生长比率的差异小于在样品内部进行两次运行之间所看到的差异。数据支持在此基础上使用p3对比野生型的成比例生长。
为评估传代次数的影响,针对每个样品绘制了计数从p-2到p0(相对于野生型)的成比例下降与所述运行中的生物体的传代次数的关系。图9示出了清晰的关系。
在此图的基础上,使用回归分析定量地评估了传代次数的影响。结果示于图10中。回归方程显示近似的线性响应,并且表明在32个传代中,各个结果的95%预测区间为相对响应10(一个数量级的差异)。基于此分析,建议最多使用32个传代,以保证相对响应(p0结果相对于p-2的比例差异)保持在10以上。
为建立p1时间点的效用,对图3中的各个曲线中的每一个曲线采取以下步骤:(1)将所有计数均转换为Log10标度;(2)利用p0、p1和p3处的响应计算斜率;这代表使用所有三个时间点时,每小时计数的变化程度;这在x轴上示出;(3)计算p3和p0处的响应的差,并除以3,以代表每小时变化;这在y轴上示出。
图11中绘制了每条曲线的两个所得变量之间的关系。所述图显示,无论是使用简单差异还是使用所有三个时间点计算的斜率,Log10(计数)中的每小时变化的值都基本相同。p1时间点在计算中不是必需的。
基于对数据的评估,支持以下内容。可以采用p3对比p0代替p5对比p0来评估样品对比野生型的相对响应。测试可以在p3处终止。传代的效果可能很显著,并且可以建议对THP-1的传代次数应用32的上限。将此上限应用于传代次数将提供从p-2到p0(野生型到样品)的响应的基线变化至少保持10倍的信心,建议将其作为下限。在p1处获得的结果对于计算样品或野生型的变化程度不是必需的。
实例4.基于THP-1的单核细胞增生性李斯特菌的感染性测定法。
ADXS11-001是一种癌症免疫治疗产品,其是经基因修饰以表达李斯特菌溶血素O(LLO)和人乳头瘤病毒(HPV)蛋白E7的融合蛋白的单核细胞增生性李斯特菌活减毒株,是一种主要发现于子宫颈癌的细胞中,但也发现于与人乳头瘤病毒16和18以及31和45直接相关联的外阴癌、阴道癌、阴茎癌和肛门癌以及口咽癌的细胞中的肿瘤抗原。
作为一种病原体,单核细胞增生性李斯特菌是一种细胞内病原体,其在被摄取到吞噬体中后逃逸进入细胞质,从而感染非吞噬细胞和吞噬细胞。这是通过表达蛋白李斯特菌溶血素O(LLO)来实现的,所述表达有助于在吞噬体与溶酶体融合以形成吞噬溶酶体之前打破液泡膜。这使得细菌能够逃逸到细胞质中,在细胞质中增殖并且直接在细胞之间扩散。THP-1细胞是人类巨噬细胞系,其在培养物中作为单核细胞保持,但可以用佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA)刺激而轻易分化为巨噬细胞。
本实例中描述的方法用于确定单核细胞增生性李斯特菌药物产品(例如,ADXS11-001)在感染分化的THP-1细胞后的离散时间点进入并且逃逸到细胞质中。以1:1的感染复数(M01)分别用野生型对照和药物产品ADXS11-001接种PMA分化的THP-1细胞。然后用庆大霉素处理感染的THP-1细胞,以杀死细胞外细菌。通过裂解THP-1细胞并且将细菌稀释液平板接种在脑心浸液(BHI)琼脂板上,在具体时间点对细菌进行感染前和感染后定量。菌落形成单位(CFU)表示由于从溶酶体中逃逸而在巨噬细胞细胞内环境中存活的活生物体。
下文示出了示例性测定法。然而,所述测定法可以用于任何李斯特菌菌株。每个测定场合可以对比对照和参考标准品评估多达2种药物产品样品。
表15.测定设置。

设备、试剂和消耗品
每个测定场合可以对比对照和参考标准品评估多达2种药物产品样品。
表16.设备。
| 设备等级 | 要求 |
| 生物安全柜 | 等级II |
| 孵育箱 | 37±1℃和5%±1CO<sub>2</sub>环境 |
| 孵育箱 | 37±1℃无CO<sub>2</sub>环境 |
| 移液管 | 2-1000μL |
| 水浴 | 由认证的温度计控制,设置为37℃ |
| 冷藏 | 2-8℃、-20℃、-70℃、-80℃和LN<sub>2</sub> |
| 离心机 | N/A |
| 微型离心机 | 1.5/2.0mL Eppendorf管,~14,500RCF |
| 涡旋 | N/A |
表17.试剂。

试剂制备指导
注:可以根据需要缩放体积/数量。
用于常规传代培养的完整RPMI(c-RPMI)(500mL):445mL RPMI 1640、50mL FBS、5mL L-谷氨酰胺(200mM)。在2-8℃下储存多达1个月。
用于解冻的完整RRMI(c-RPMI-解冻)505mL:400mL RPMI 1640、100mL FBS、5mL L-谷氨酰胺(200mM),在2-8℃下储存多达1个月。
冷冻溶液500mL:新鲜制备的450mL热灭活FBS、50mL甘油。
1.6mM PMA:1.0mg PMA(Mw:616.83)、1.0mL DMSO,在-20℃下储存多达6个月。在2mL无菌微型离心管中制备10μL等分试样。每个等分试样都是一次性使用的。
100mg/mL链霉素:1g链霉素、10mL无菌水,使用0.2μm过滤器消毒,并且在-20℃下储存多达1个月。在2mL无菌微型离心管中制备1mL等分试样。每个等分试样都是一次性使用的。
脑心浸液琼脂+100μg/mL链霉素。在进行之前对BHI板进行检验,以验证其是否存在制造缺陷(污染、断板、琼脂不均匀等)。每个琼脂板的体积为大约22.8mL。177.2μL×琼脂板数PBS、22.8μL×琼脂板数100mg/mL、链霉素100mg/mL。向每个琼脂板中加入200μL稀释的链霉素。使用无菌涂布器涂布以覆盖板的整个表面。涂布直到琼脂板吸收所有液体。将板储存在2-8℃的温度下直到琼脂板或链霉素中较早者的有效期。
THP-1细胞系培养
解冻THP-1细胞。在无菌条件下在生物安全柜中执行程序。仅使用经过认证的无菌材料和无菌制备的材料。
1.在设置为37℃的水浴中预热c-RPMI-解冻培养基。
2.将3mL预热的c-RPMI-解冻培养基放入无菌的50mL离心管中。
3.从低温储存器中取出THP-1小瓶,并且在设置为37℃的水浴中解冻,直到内容物几乎解冻,但仍有少量冰晶在管中。
4.用消毒剂彻底清洁小瓶。
5.将解冻的细胞逐滴加入含有3mL c-RPMI-解冻培养基的50mL离心管中。
6.用另外的1mL c-RPMI-解冻培养基清洗冷冻瓶,并将其转移到含有细胞的50mL管中。
7.取~100μL细胞悬液进行计数。
注:用血细胞计数仪计数时,将细胞悬液等分试样1:2稀释于0.4%台盼蓝中。通过轻轻移液确保混悬液充分混合。使用C芯片进行计数。进行两次独立计数。测定细胞生存力(≥85%)和密度。
8.在室温下以150×g离心细胞悬液,持续5分钟
9.丢弃上清液,将细胞重新悬浮在预热的c-RPMI-解冻培养基中,使细胞密度达到1-3×105个活细胞/mL。
10.将管中的内容物转移到细胞培养瓶(例如T75),并且在37℃5%CO2绝对湿度下孵育。
11.将烧瓶保持在竖直位置,直到细胞达到指数生长期。
12.通常每2-3天计数一次细胞。
注:一旦建立了培养物(通常在解冻6天后),则使用c-RPMI培养基将血清浓度降低到10%。
常规THP-1细胞培养。使用经过认证的无菌材料和无菌制备的材料在无菌条件下在生物安全柜中执行程序。
在一些实施例中,对于常规细胞培养和THP-1测定,细胞传代次数限于P32。细胞每次转移到一个新的培养皿中被认为是一次传代。向同一培养皿中加入培养基以确保指数生长不会改变传代次数。
为使细胞保持指数生长,将培养物保持在3-8×105个活细胞/mL之间。
1.在显微镜下检查细胞的形态和污染。
如果大多数细胞附着在培养皿表面,则勿继续操作。在这种情况下,丢弃培养物并且解冻另一瓶工作细胞库。
2.将约1mL细胞悬液移入小瓶中以测定总细胞计数和生存力。
3.为使细胞处于指数生长期,向细胞中将补充新鲜的c-RPMI培养基,使其密度达到3×105个细胞/mL,直到细胞悬液体积达到最大允许体积,然后将细胞悬液以3×105个细胞/mL的接种密度传代到新的预标记烧瓶中。
下面显示了不同大小烧瓶的最小和最大体积范围,以实现最佳的CO2渗透:775烧瓶:15-37.5mL;T150烧瓶:30-75mL
4.在37℃的5%CO2孵育箱中孵育培养物。
5.通常每2-3天计数一次细胞。
THP-1细胞的冷冻保存。使用经过认证的无菌材料和无菌制备的材料在无菌条件下在生物安全柜中执行程序。
1.遵循“常规THP-1细胞培养”步骤1到2。
2.制备由补充有10%(v/v)甘油的热灭活FBS组成的冷冻培养基。
3.在室温下以150×g离心细胞,持续5分钟。
4.丢弃上清液,并通过轻敲管使细胞重新悬浮,直到看不到团块。通过旋转管缓慢地逐滴添加冷冻培养基,从而得到2倍最终冷冻密度(最终冷冻密度为2×106个细胞/mL)。
5.向含有细胞的管中缓慢地添加第二个等体积的冷冻培养基。在添加期间轻轻地旋转试管使其完全混合。
6.用血清学移液管将1mL细胞悬液等分放入预先标记的2mL冷冻瓶中。
7.将冷冻瓶放入室温的CoolCell或Mr Frosty容器中,用2-丙醇填充到标记处。
8.将冷冻容器转移到-70℃的冰箱中,持续24-72小时。
9.将冷冻瓶转移到气相氮气储存器中。
10.记录冷冻细胞批次的位置和详细信息。
THP-1细胞的制备和细胞分化(第1天)注:每个测试项目(对照、参考品或样品)准备l×THP-1 24孔板。每个板至少需要7个经PMA处理的孔和2个未经处理的孔。示例板布局如下所示。
表18. 24孔板。
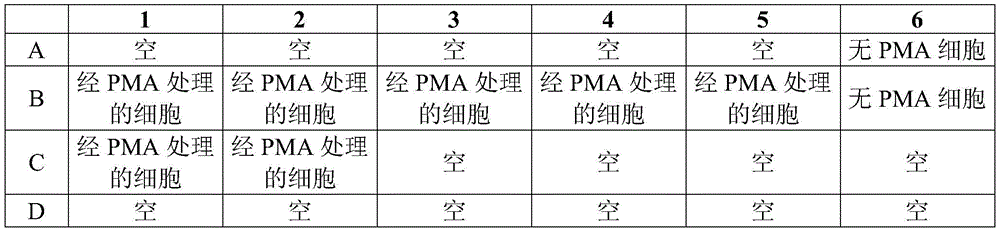
板布局包含应急孔。
1.在设置为37℃的水浴中预热完整的RPMI(c-RPMI)培养基。
2.从孵育箱中去除细胞,目视检查是否有污染迹象,并在显微镜下检查细胞。
如果污染,勿继续操作。如果大多数细胞附着在培养皿表面,勿继续操作。在这种情况下,丢弃培养物并且解冻另一瓶工作细胞库。
3.将细胞悬液上下移液几次,以混合细胞,并取出少量细胞进行细胞计数。
4.在0.4%台盼蓝中制备细胞悬液的1/2稀释液。通过用移液管轻轻混合,确保稀释的细胞悬液正确混合。
5.为总共2个独立的细胞计数准备C芯片,并确定细胞密度和生存力。仅在细胞生存力为至少85%时继续操作。
6.以1×106个活细胞/mL制备细胞悬液:在室温下以150×g离心适当体积的细胞悬液,持续5分钟,丢弃上清液并将细胞沉淀重新悬浮于c-RPMI中。均匀混合。每孔至少准备1mL细胞悬液。
7.向24孔板的标有“NO PMA”的两个孔中的每一个中分别添加1mL细胞悬液(参见板布局)。
8.向剩余的细胞悬液中添加16pM(来自于1.6mM储备液中的1/100稀释液)PMA,直到最终浓度为大约16nM PMA。均匀混合。
9.将每孔1mL经PMA处理的细胞悬液添加到每块板上的至少7孔中(参见板布局)。
10.在37±1℃、5±1%CO2条件下孵育细胞,持续16-24小时。
THP-1细胞的感染和时间进程(第2天)
样品和对照的制备。对含有经PMA处理的THP-1细胞的孔的所有操作应当小心处理。应当通过将板倾斜大约45度角来抽吸或分配培养基。抽吸或分配步骤中,移液管尖端不得擦伤孔表面。
1.取出一小瓶野生型单核细胞增生性李斯特菌/参考标准品或药物产品样品。
2.室温下解冻小瓶持续多达10分钟,并确保样品完全解冻。
3.涡旋并将细胞悬液转移到分别标记的2mL微量离心管(1mL)中。记录准确的转移量。
4.在室温下以14500×g离心细胞,持续2分钟
5.小心地丢弃上清液,并用RT-c-RPMI重新悬浮沉淀物。培养基的体积应当等于步骤3中最初转移的测试项目的体积。
6.使用c-RPMI制备细菌稀释液直到最终浓度为1.0×106CFU/mL。此浓度下的最终体积应为大约15mL。
由于单核细胞增生性李斯特菌可以在c-RPMI中生长,请立即进入下一章节
用单核细胞增生性李斯特菌感染THP-1细胞。
1.从孵育箱中去除一个24孔板。
2.在显微镜下确认THP-1细胞的粘附性并且确认经PMA处理的细胞(轻度摇动时仍保持粘附)与未经处理的细胞(轻度摇动时呈现出流动性)之间的区别。
3.从含有经PMA处理的细胞的孔中吸出培养基并且添加1mL在“样品和对照的制备”的步骤7中制备的细菌。
4.在显微镜上观察板以确保THP-1细胞仍粘附在孔表面。
5.在37±1℃、5±1%CO2下孵育板,持续2小时±3分钟。
6.按照“生存力测试程序—p-2时间点”执行在“样品和对照的制备”中制备的测试项目的生存力测试。
生存力测试程序—p-2时间点。在启动生存力测试之前确保所有琼脂板都充分干燥。
1.使用按照描述稀释至1.0×106CFU/mL的测试项目(THP 1细胞的感染和时间进程(第2天))。
2.按照下表顺序稀释细菌悬液:

3.将100μL的每种稀释液涂布在适当的BHI琼脂板上。针对每种稀释液制作3个琼脂板(即,对于p-2时间点,每个测试项目共产生9个琼脂板)。
将琼脂BHI板用于野生型单核细胞增生性李斯特菌对照
将琼脂BM+氯霉素用于ADXS11-001测试项目
4.使每个板吸收液体并盖上盖子至少干燥15分钟,然后将其倒置并且放置在35-38℃的无CO2的孵育箱中,持续16-24小时。
停止感染。
1.制备含有20pg/mL庆大霉素的c-RPMI。
2. 2小时后,从孵育箱中去除含有野生型或样品的板。
3.使用移液管从每个孔中去除含有单核细胞增生性李斯特菌的培养基。
4.每孔小心分配1mL制备好的含有20pg/mL庆大霉素的c-RPMI,向孔的一侧缓慢地添加以避免破坏。
5.将板放回37±1℃、5±1%CO2的孵育箱中,持续45分钟。
6.使用移液管从每个孔中去除含有20pg/ml庆大霉素的c-RPMI。
7.通过在每孔中加入1mL不含庆大霉素的c-RPMI(向孔的一侧缓慢地添加以避免单层破坏),小心地洗涤细胞。
8.使用移液管从每个孔中去除c-RMPI。
9.通过向孔的一侧缓慢地添加以避免破坏而小心地将1mL的c-RPMI(不含庆大霉素)分配到每个孔中,并且将板放回设置为37±1℃、5±1%CO2的孵育箱中,持续至少5分钟。孵育结束时间指定为p0。
细胞内单核细胞增生性李斯特菌生长的检测—p0。
1.选择一个孔用于时间点“p0”的收集。
确保每个孔中经PMA处理的THP-1细胞的层是一致的,并且确保在之前的抽吸和分配步骤期间很少或没有细胞被移出和去除。如果观察到任何孔有显著的THP-1细胞丢失,则做好标记以提示不要使用这些孔。
2.通过用移液管抽吸,从所选的孔中去除c-RPMI。
3.向孔中分配1mL无菌水。通过上下移液将THP-1细胞从孔表面移出。将全部内容物转移到2mL离心管中
在显微镜下观察以确认细胞已成功去除。如果仍然残留大量THP-1细胞,则利用先前转移到2mL管中的一部分水通过上下移液移出细胞。将内容物转移回2mL管中,并在显微镜下确认THP-1细胞已被去除。
4.将板放回37±1℃、5±1%CO2的孵育箱中直到准备好收集下一个时间点为止
5.涡旋细胞裂解物至少1分钟以释放细胞内细菌,并且按照“生存力测试程序—p0/p3时间点”进行生存力测试。
细胞内单核细胞增生性李斯特菌生长的检测—p3。
1.在指定时间p0后3小时后从孵育箱中取出板(“停止感染”,步骤9)。
2.选择一个孔用于时间点“p3”的采集。
确保每个孔中经PMA处理的THP-1细胞的层是一致的,并且确保在之前的抽吸和分配步骤期间很少或没有细胞被移出和去除。如果观察到任何孔有显著的THP-1细胞丢失,则做好标记以提示不要使用这些孔。
3.通过用移液管抽吸,从所选的孔中去除c-RPM1。
4.向孔中分配1mL无菌水。通过上下移液将THP-1细胞从孔表面移出。将全部内容物转移到2mL离心管中。
在显微镜下观察孔以确认细胞已成功去除。如果仍然残留大量THP-1细胞,则利用先前转移到2mL管中的一部分水通过上下移液移出细胞。将内容物转移回2mL管中,并在显微镜下确认THP-1细胞已被去除。
5.涡旋细胞裂解物至少1分钟以释放细胞内细菌,并且按照“生存力测试程序—p0/p3时间点”进行生存力测试。
生存力测试程序—p0/p3时间点。
1.使用在以下步骤中生成的测试项目裂解物:
对于p0:“细胞内单核细胞增生性李斯特菌生长的检测—p0,步骤5”
对于p3:“细胞内单核细胞增生性李斯特菌生长的检测—p3,步骤5”
2.按照下表顺序稀释细菌悬液:

3.将100μL的每种稀释液涂布在适当的BHI琼脂板上。针对每种稀释液制作3个琼脂板(即,对于p-2时间点,每个测试项目共产生9个琼脂板)。
将琼脂BHI板用于野生型单核细胞增生性李斯特菌对照。
将琼脂BHI+氯霉素用于ADXS11-001测试项目。
4.将使每个板吸收液体并盖上盖子至少干燥15分钟,然后将其倒置并且放置在35-38℃的无CO2的孵育箱中,持续16-24小时。
对照板。
1.对于每种测定场合,准备以下阴性对照琼脂板:
未接种的:
3×BHI琼脂+100pg/mL链霉素
3×BHI琼脂+25pg/mL氯霉素
接种的:
接种了100μL PBS的3×BHI琼脂+100pg/mL链霉素
接种了100μL PBS的3×BHI琼脂+25pg/mL氯霉素
2.对于每种测定场合,准备以下阳性对照琼脂板:
以1×106CFU/mL接种了10μL野生型单核细胞增生性李斯特菌的2×BHI琼脂+100pg/mL链霉素。
以1×106CFU/mL接种了10μL参考标准品的2×BHI琼脂+25pg/mL氯霉素。
3.将板与测定琼脂板一起在35-38℃(无CO2)下孵育16-24小时。
菌落计数(第3天)
1.16-24小时后,从孵育箱中取出板
注:确保在每个时间点将所有单核细胞增生性李斯特菌细胞类型(野生型、参考标准品和样品)孵育相同的持续时间。
2.将针对每种稀释液的每个板手动计数并在工作表中记录菌落形成单位的总数。
计算
1.仅将值在40-600之间的菌落计数用于后续计算。每种稀释液至少要有2个菌落计数在范围内才能进行必要的计算。如果两个以上的板的菌落计数在40-600范围外,则在p0和/或p3时间点使用调整后的稀释液重复整个测定(“生存力测试程序—p0/p3时间点”,步骤2)。
2.针对每个时间点计算CFU/mL值:

3.对所有计算的CFU/mL值进行log10转换。
4.在y轴上用log10 CFU/mL绘制数据,并在x轴上绘制时间。
测定接受标准
1.所有阴性对照琼脂板上均无细菌生长的迹象。
2.所有阳性对照琼脂板上均应存在菌落。
3.在p-2处计算的对照的平均log10(CFU/mL)在6±0.5以内
4.一式三份的琼脂板的有效菌落计数值之间的%CV*≤30。
*CV=[(标准差/平均值)×100]
5.根据以下公式计算细胞系性能(CLP)参数:

对于所有有效的测定运行,CLP≥3。CLP值需要跟踪。P-2、p0=分别针对时间点p-2和p0的平均CFU/mL值。
6.使用以下公式计算对照的参考响应:

对于所有有效的测定运行,RRS≥2.0。RRS值需要跟踪。P3、p0=分别针对时间点p3和p0的平均CFU/mL值。
可报告结果
1.针对每个样品使用以下公式计算可报告结果:

报告到小数点后1位(d.p.)。
2.根据规范评估结果。
实例5.基于THP-1的单核细胞增生性李斯特菌的感染性测定法的验证。
实例4提供了所述方法的概述。
表19.总结。

方法学
测定需要3天完成。第1天,将THP-1细胞以1×106个活细胞/mL平板接种在24孔组织培养板中(每个测试项目一块板—参见上文(THP 1细胞的感染和时间进程(第2天))。仅使用生存力大于85%的细胞,并且培养物的传代次数限于P32。然后将THP-1细胞用PMA溶液处理,以在过夜孵育期间刺激其分化为巨噬细胞。
第二天使用光学显微镜通过目视确认分化。分化的细胞粘附于孔表面并且在形态上与保留在混悬液中的未分化的圆形细胞不同。
将每个测试项目的浓度调整至1×106CFU/mL(基于标称浓度),并且进一步连续稀释至10-1、10-2和10-3。将100μL的每种稀释液平板接种在BHI琼脂板上并孵育16-24小时,以使菌落生长。针对每种稀释液制备3个板。然后手动计数菌落,以产生p-2CFU/mL值。在这个时间点,感染前的固定CFU/mL量预计将产生6±0.5log10 CFU/mL。这确保使用相同数量的测试项目来感染THP-1细胞。
还将调整至1×106CFU/mL的测试项目添加到分化的THP-1细胞中,持续2小时±3分钟。在此期间,单核细胞增生性李斯特菌细菌进入THP-1细胞。然后通过添加庆大霉素持续45分钟,杀死培养基中残留的所有细菌。庆大霉素无法穿透THP-1细胞的细胞膜,并且因此在此步骤中仅去除了细胞外细菌。将含有单核细胞增生性李斯特菌的THP-1细胞裂解。将裂解物连续稀释至10-1、10-2和10-3。将100μL的每种稀释液平板接种在BHI琼脂板上并孵育16-24小时,以使菌落生长。针对每种稀释液制备3个板。然后手动计数菌落,以产生p0CFU/mL值。此时,确定每个测试项目的感染细菌细胞数。
用庆大霉素完成处理后,将含有感染单核细胞增生性李斯特菌的THP-1细胞的几个孔留在孵育箱中,持续3小时。在孵育时间结束时裂解细胞。将裂解物连续稀释至10-1、10-2和10-3。将100μL的每种稀释液平板接种在BHI琼脂板上并孵育16-24小时,以使菌落生长。针对每种稀释液制备3个板。然后手动计数菌落,以产生p3 CFU/mL值。此时,确定每个测试项目的感染进展。
还准备了对照板以通过抗生素抗性曲线评估无菌技术以及测试项目的同一性。将对照板与p-2、p0和p3 BHI琼脂板一起孵育。
数据分析
手动计数每个BHI琼脂板。每个菌落等于1CFU。每种调配物/裂解物稀释液(即10-1、10-2和10-3)给出3个菌落计数值(即CFU)。预计至少一种稀释液将产生40-600个菌落/BHI琼脂板内的菌落计数,并且%CV<30%。根据以下方程计算每个测试项目在p-2、p0和p3时间点处的CFU/mL值:

对于所有有效的测定运行,预计对照在p-2处的log10(CFU/mL)在6±0.5以内。
为评估每个测试项目的细胞内生长,使用以下方程计算可报告结果:

其中p3、p0=分别针对时间点p3和p0的平均CFU/mL。
可报告结果计算到小数点后1位。
预计参考标准材料的可报告结果≥2.0。
另外,通过计算细胞系性能参数来测量分化的THP-1细胞对感染的容许性:

其中p-2、p0=分别针对时间点P-2和p0的平均CFU/mL。
细胞系性能参数计算到小数点后0位。
对于所有有效的测定运行,预计细胞系参数≥3。
方法性能参数的评估方法
表20.分析矩阵。

Ref.=参考(reference)
测定内精度。对于测定内精度,数据是从一种测定场合(A1)收集的,所述测定场合由一式三份(n=3)进行测试的参考材料和一个对照(n=1)组成。数据反映了在相同分析条件下的可变性。制备参考材料(n=3)调配物,并且在相同的测定场合进行独立处理。
计算:参考标准品(测定A1-A7中的测试项目2)的平均值/SD可报告结果+%CV;n=7。
特异性。测定的特异性被定义为测试系统区分对照与参考材料/样品的生长模式的能力。
为考虑时间和项目对CFU/mL的影响,以项目、时间和其相互作用作为固定因素并且将包含的重复项作为随机影响进行了双向方差分析(ANOVA)。相互作用影响描述了每个项目在时间进程上的差异。分析前对数据进行对数转换(以10为底)。
接着上述ANOVA,使用双单侧测试方法(TOST)将每个项目的等效性与对照进行比较。对于每个比较,确定对照平均值与项目平均值之间差异的置信区间。考虑均值之间的差异的等效区间(-0.5,0.5),确定两个均值之间的差异的90%置信区间。如果两个置信限都在等效区间内,则这两个均值被声明为等效。
计算由ENVIGO统计部门使用SAS软件(使用Proc GLM的9.1.3版)进行。
鲁棒性确认。根据预验证研究的结果,THP-1细胞的感染时间被限定为2小时+/-3分钟。为证明此范围对可报告结果无影响,使用感染时间的下限和上限(A6和A7)进行了2次测定。将测定A1(n=3)的平均可报告结果与测定A6(n=3)和A7(n=3)的平均可报告结果进行比较。预计A1、A6和A7的%CV≤25。
表21.关键材料。
| 材料 | 供应商 | 标称浓度 | 批次 |
| ADXS11-001(参考材料) | Advaxis公司 | 8.8×10<sup>9</sup>CFU/mL | 5265-14-01 |
| 野生型单核细胞增生性李斯特菌(对照) | Advaxis公司 | 1.7×10<sup>9</sup>CFU/mL | NB89p25 |
结果
表22.测定接受标准评估。

1CLP报告为具有多于一个参考测试项目的测定的平均值。
210-2稀释液和10-3稀释液各自的%CV。
3RRS报告为具有多于一个参考测试项目的测定的平均值。
表23.测定内精度。

表24.测定内精度。

1鲁棒性确认。测定A6评估了增加的感染时间(2小时±3分钟)
表25.测定内精度。

1鲁棒性确认。测定A7评估了减少的感染时间(2小时±3分钟)。
表26.测定间精度(中间)。

表27.特异性。
| 对比 | 时间点 | 差值 | 下限CL | 上限CL | 相等 |
| 项目2与对照 | -2 | -0.12 | -0.18 | -0.07 | 是 |
| 项目2与对照 | 0 | -1.55 | -1.60 | -1.51 | 否 |
| 项目2与对照 | 3 | -2.18 | -2.23 | -2.13 | 否 |
| 项目3与对照 | -2 | -0.14 | -0.19 | -0.10 | 是 |
| 项目3与对照 | 0 | -1.68 | -1.73 | -1.64 | 否 |
| 项目3与对照 | 3 | -2.28 | -2.32 | -2.23 | 否 |
| 项目4与对照 | -2 | -0.10 | -0.15 | -0.06 | 是 |
| 项目4与对照 | 0 | -1.62 | -1.67 | -1.57 | 否 |
| 项目4与对照 | 3 | -2.28 | -2.32 | -2.23 | 否 |
表28.

表29.菌落计数。

1–菌落计数超出允许范围(40-600个菌落)。
序列表
<110> 阿德瓦希斯公司
帕纳姆·莫利
安努·瓦勒查
<120> 用于评估李斯特菌菌株的减毒和感染性的组合物和方法
<130> 062384/528092
<150> 62/640,855
<151> 2018-03-09
<160> 99
<170> PatentIn版本3.5
<210> 1
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 1
gcacgtagta taatcaactt tgaaaaactg taataa 36
<210> 2
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 2
gcacgttcta ttatcaactt cgaaaaacta taataa 36
<210> 3
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 3
gcccgcagta ttatcaattt cgaaaaatta taataa 36
<210> 4
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 4
gcgcgctcta taattaactt cgaaaaactt taataa 36
<210> 5
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 5
gcacgctcca ttattaactt tgaaaaactt taataa 36
<210> 6
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 6
gctcgctcta tcatcaattt cgaaaaactt taataa 36
<210> 7
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 7
gcacgtagta ttattaactt cgaaaagtta taataa 36
<210> 8
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 8
gcacgttcca tcattaactt tgaaaaacta taataa 36
<210> 9
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 9
gctcgctcaa tcatcaactt tgaaaagcta taataa 36
<210> 10
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 10
gctcgctcta tcatcaactt cgaaaaattg taataa 36
<210> 11
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 11
gctcgctcta ttatcaattt tgaaaaatta taataa 36
<210> 12
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 12
gctcgtagta ttattaattt cgaaaaatta taataa 36
<210> 13
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 13
gctcgttcga ttatcaactt cgaaaaactg taataa 36
<210> 14
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 14
gcaagaagca tcatcaactt cgaaaaactg taataa 36
<210> 15
<211> 36
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 15
gcgcgttcta ttattaattt tgaaaaatta taataa 36
<210> 16
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 16
Ala Arg Ser Ile Ile Asn Phe Glu Lys Leu
1 5 10
<210> 17
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 17
gattataaag atcatgacgg agactataaa gaccatgaca ttgattacaa agacgacgat 60
gacaaa 66
<210> 18
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 18
gactataaag accacgatgg cgattataaa gaccatgata ttgactacaa agatgatgat 60
gataag 66
<210> 19
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 19
gattataaag atcatgatgg cgactataaa gatcatgata tcgattacaa agatgacgat 60
gacaaa 66
<210> 20
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 20
gactacaaag atcacgatgg tgactacaaa gatcacgaca ttgattataa agacgatgat 60
gacaaa 66
<210> 21
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 21
gattacaaag atcacgatgg tgattataag gatcacgata ttgattacaa agacgacgac 60
gataaa 66
<210> 22
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 22
gattacaaag atcacgatgg cgattacaaa gatcatgaca ttgactacaa agacgatgat 60
gataaa 66
<210> 23
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 23
gattacaagg atcatgatgg tgattacaaa gatcacgata tcgactacaa agatgatgac 60
gataaa 66
<210> 24
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 24
gactacaaag atcatgatgg tgattacaaa gatcatgaca ttgattataa agatgatgat 60
gacaaa 66
<210> 25
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 25
gattataaag accatgatgg tgattataag gatcatgata tcgattataa ggatgacgac 60
gataaa 66
<210> 26
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 26
gattataaag atcacgatgg cgattataaa gaccacgata ttgattataa agacgacgat 60
gacaaa 66
<210> 27
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 27
gactataaag accacgatgg tgattataaa gatcacgaca tcgactacaa agacgatgat 60
gataaa 66
<210> 28
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 28
gactacaaag atcacgacgg cgattataaa gatcacgata ttgactataa agatgacgat 60
gataaa 66
<210> 29
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 29
gattataaag accatgatgg agattacaaa gatcatgata ttgactataa agacgacgac 60
gataaa 66
<210> 30
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 30
gattataaag atcacgatgg tgactacaaa gatcacgata tcgattataa agacgatgac 60
gataaa 66
<210> 31
<211> 66
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 31
gactacaaag atcacgatgg tgattataaa gaccatgata ttgattacaa agatgatgat 60
gacaaa 66
<210> 32
<211> 22
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 32
Asp Tyr Lys Asp His Asp Gly Asp Tyr Lys Asp His Asp Ile Asp Tyr
1 5 10 15
Lys Asp Asp Asp Asp Lys
20
<210> 33
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 33
Gly Ala Ser Gly Ala Ser
1 5
<210> 34
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 34
Gly Ser Ala Gly Ser Ala
1 5
<210> 35
<211> 4
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 35
Gly Gly Gly Gly
1
<210> 36
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 36
Gly Gly Gly Gly Ser
1 5
<210> 37
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 37
Val Gly Lys Gly Gly Ser Gly Gly
1 5
<210> 38
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 38
Pro Ala Pro Ala Pro
1 5
<210> 39
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 39
Glu Ala Ala Ala Lys
1 5
<210> 40
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 40
Ala Tyr Leu Ala Tyr Leu
1 5
<210> 41
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 41
Leu Arg Ala Leu Arg Ala
1 5
<210> 42
<211> 4
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 42
Arg Leu Arg Ala
1
<210> 43
<211> 32
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 43
Lys Glu Asn Ser Ile Ser Ser Met Ala Pro Pro Ala Ser Pro Pro Ala
1 5 10 15
Ser Pro Lys Thr Pro Ile Glu Lys Lys His Ala Asp Glu Ile Asp Lys
20 25 30
<210> 44
<211> 19
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 44
Lys Glu Asn Ser Ile Ser Ser Met Ala Pro Pro Ala Ser Pro Pro Ala
1 5 10 15
Ser Pro Lys
<210> 45
<211> 14
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 45
Lys Thr Glu Glu Gln Pro Ser Glu Val Asn Thr Gly Pro Arg
1 5 10
<210> 46
<211> 28
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 46
Lys Glu Ser Val Val Asp Ala Ser Glu Ser Asp Leu Asp Ser Ser Met
1 5 10 15
Gln Ser Ala Asp Glu Ser Thr Pro Gln Pro Leu Lys
20 25
<210> 47
<211> 20
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 47
Lys Ser Glu Glu Val Asn Ala Ser Asp Phe Pro Pro Pro Pro Thr Asp
1 5 10 15
Glu Glu Leu Arg
20
<210> 48
<211> 33
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 48
Arg Gly Gly Arg Pro Thr Ser Glu Glu Phe Ser Ser Leu Asn Ser Gly
1 5 10 15
Asp Phe Thr Asp Asp Glu Asn Ser Glu Thr Thr Glu Glu Glu Ile Asp
20 25 30
Arg
<210> 49
<211> 17
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 49
Lys Gln Asn Thr Ala Ser Thr Glu Thr Thr Thr Thr Asn Glu Gln Pro
1 5 10 15
Lys
<210> 50
<211> 17
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 50
Lys Gln Asn Thr Ala Asn Thr Glu Thr Thr Thr Thr Asn Glu Gln Pro
1 5 10 15
Lys
<210> 51
<211> 19
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 51
Arg Ser Glu Val Thr Ile Ser Pro Ala Glu Thr Pro Glu Ser Pro Pro
1 5 10 15
Ala Thr Pro
<210> 52
<211> 28
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 52
Lys Ala Ser Val Thr Asp Thr Ser Glu Gly Asp Leu Asp Ser Ser Met
1 5 10 15
Gln Ser Ala Asp Glu Ser Thr Pro Gln Pro Leu Lys
20 25
<210> 53
<211> 20
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 53
Lys Asn Glu Glu Val Asn Ala Ser Asp Phe Pro Pro Pro Pro Thr Asp
1 5 10 15
Glu Glu Leu Arg
20
<210> 54
<211> 33
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 54
Arg Gly Gly Ile Pro Thr Ser Glu Glu Phe Ser Ser Leu Asn Ser Gly
1 5 10 15
Asp Phe Thr Asp Asp Glu Asn Ser Glu Thr Thr Glu Glu Glu Ile Asp
20 25 30
Arg
<210> 55
<211> 529
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 55
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Lys Asp Ala Ser Ala Phe Asn Lys
20 25 30
Glu Asn Ser Ile Ser Ser Met Ala Pro Pro Ala Ser Pro Pro Ala Ser
35 40 45
Pro Lys Thr Pro Ile Glu Lys Lys His Ala Asp Glu Ile Asp Lys Tyr
50 55 60
Ile Gln Gly Leu Asp Tyr Asn Lys Asn Asn Val Leu Val Tyr His Gly
65 70 75 80
Asp Ala Val Thr Asn Val Pro Pro Arg Lys Gly Tyr Lys Asp Gly Asn
85 90 95
Glu Tyr Ile Val Val Glu Lys Lys Lys Lys Ser Ile Asn Gln Asn Asn
100 105 110
Ala Asp Ile Gln Val Val Asn Ala Ile Ser Ser Leu Thr Tyr Pro Gly
115 120 125
Ala Leu Val Lys Ala Asn Ser Glu Leu Val Glu Asn Gln Pro Asp Val
130 135 140
Leu Pro Val Lys Arg Asp Ser Leu Thr Leu Ser Ile Asp Leu Pro Gly
145 150 155 160
Met Thr Asn Gln Asp Asn Lys Ile Val Val Lys Asn Ala Thr Lys Ser
165 170 175
Asn Val Asn Asn Ala Val Asn Thr Leu Val Glu Arg Trp Asn Glu Lys
180 185 190
Tyr Ala Gln Ala Tyr Pro Asn Val Ser Ala Lys Ile Asp Tyr Asp Asp
195 200 205
Glu Met Ala Tyr Ser Glu Ser Gln Leu Ile Ala Lys Phe Gly Thr Ala
210 215 220
Phe Lys Ala Val Asn Asn Ser Leu Asn Val Asn Phe Gly Ala Ile Ser
225 230 235 240
Glu Gly Lys Met Gln Glu Glu Val Ile Ser Phe Lys Gln Ile Tyr Tyr
245 250 255
Asn Val Asn Val Asn Glu Pro Thr Arg Pro Ser Arg Phe Phe Gly Lys
260 265 270
Ala Val Thr Lys Glu Gln Leu Gln Ala Leu Gly Val Asn Ala Glu Asn
275 280 285
Pro Pro Ala Tyr Ile Ser Ser Val Ala Tyr Gly Arg Gln Val Tyr Leu
290 295 300
Lys Leu Ser Thr Asn Ser His Ser Thr Lys Val Lys Ala Ala Phe Asp
305 310 315 320
Ala Ala Val Ser Gly Lys Ser Val Ser Gly Asp Val Glu Leu Thr Asn
325 330 335
Ile Ile Lys Asn Ser Ser Phe Lys Ala Val Ile Tyr Gly Gly Ser Ala
340 345 350
Lys Asp Glu Val Gln Ile Ile Asp Gly Asn Leu Gly Asp Leu Arg Asp
355 360 365
Ile Leu Lys Lys Gly Ala Thr Phe Asn Arg Glu Thr Pro Gly Val Pro
370 375 380
Ile Ala Tyr Thr Thr Asn Phe Leu Lys Asp Asn Glu Leu Ala Val Ile
385 390 395 400
Lys Asn Asn Ser Glu Tyr Ile Glu Thr Thr Ser Lys Ala Tyr Thr Asp
405 410 415
Gly Lys Ile Asn Ile Asp His Ser Gly Gly Tyr Val Ala Gln Phe Asn
420 425 430
Ile Ser Trp Asp Glu Val Asn Tyr Asp Pro Glu Gly Asn Glu Ile Val
435 440 445
Gln His Lys Asn Trp Ser Glu Asn Asn Lys Ser Lys Leu Ala His Phe
450 455 460
Thr Ser Ser Ile Tyr Leu Pro Gly Asn Ala Arg Asn Ile Asn Val Tyr
465 470 475 480
Ala Lys Glu Cys Thr Gly Leu Ala Trp Glu Trp Trp Arg Thr Val Ile
485 490 495
Asp Asp Arg Asn Leu Pro Leu Val Lys Asn Arg Asn Ile Ser Ile Trp
500 505 510
Gly Thr Thr Leu Tyr Pro Lys Tyr Ser Asn Lys Val Asp Asn Pro Ile
515 520 525
Glu
<210> 56
<211> 529
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 56
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Lys Asp Ala Ser Ala Phe Asn Lys
20 25 30
Glu Asn Ser Ile Ser Ser Val Ala Pro Pro Ala Ser Pro Pro Ala Ser
35 40 45
Pro Lys Thr Pro Ile Glu Lys Lys His Ala Asp Glu Ile Asp Lys Tyr
50 55 60
Ile Gln Gly Leu Asp Tyr Asn Lys Asn Asn Val Leu Val Tyr His Gly
65 70 75 80
Asp Ala Val Thr Asn Val Pro Pro Arg Lys Gly Tyr Lys Asp Gly Asn
85 90 95
Glu Tyr Ile Val Val Glu Lys Lys Lys Lys Ser Ile Asn Gln Asn Asn
100 105 110
Ala Asp Ile Gln Val Val Asn Ala Ile Ser Ser Leu Thr Tyr Pro Gly
115 120 125
Ala Leu Val Lys Ala Asn Ser Glu Leu Val Glu Asn Gln Pro Asp Val
130 135 140
Leu Pro Val Lys Arg Asp Ser Leu Thr Leu Ser Ile Asp Leu Pro Gly
145 150 155 160
Met Thr Asn Gln Asp Asn Lys Ile Val Val Lys Asn Ala Thr Lys Ser
165 170 175
Asn Val Asn Asn Ala Val Asn Thr Leu Val Glu Arg Trp Asn Glu Lys
180 185 190
Tyr Ala Gln Ala Tyr Ser Asn Val Ser Ala Lys Ile Asp Tyr Asp Asp
195 200 205
Glu Met Ala Tyr Ser Glu Ser Gln Leu Ile Ala Lys Phe Gly Thr Ala
210 215 220
Phe Lys Ala Val Asn Asn Ser Leu Asn Val Asn Phe Gly Ala Ile Ser
225 230 235 240
Glu Gly Lys Met Gln Glu Glu Val Ile Ser Phe Lys Gln Ile Tyr Tyr
245 250 255
Asn Val Asn Val Asn Glu Pro Thr Arg Pro Ser Arg Phe Phe Gly Lys
260 265 270
Ala Val Thr Lys Glu Gln Leu Gln Ala Leu Gly Val Asn Ala Glu Asn
275 280 285
Pro Pro Ala Tyr Ile Ser Ser Val Ala Tyr Gly Arg Gln Val Tyr Leu
290 295 300
Lys Leu Ser Thr Asn Ser His Ser Thr Lys Val Lys Ala Ala Phe Asp
305 310 315 320
Ala Ala Val Ser Gly Lys Ser Val Ser Gly Asp Val Glu Leu Thr Asn
325 330 335
Ile Ile Lys Asn Ser Ser Phe Lys Ala Val Ile Tyr Gly Gly Ser Ala
340 345 350
Lys Asp Glu Val Gln Ile Ile Asp Gly Asn Leu Gly Asp Leu Arg Asp
355 360 365
Ile Leu Lys Lys Gly Ala Thr Phe Asn Arg Glu Thr Pro Gly Val Pro
370 375 380
Ile Ala Tyr Thr Thr Asn Phe Leu Lys Asp Asn Glu Leu Ala Val Ile
385 390 395 400
Lys Asn Asn Ser Glu Tyr Ile Glu Thr Thr Ser Lys Ala Tyr Thr Asp
405 410 415
Gly Lys Ile Asn Ile Asp His Ser Gly Gly Tyr Val Ala Gln Phe Asn
420 425 430
Ile Ser Trp Asp Glu Val Asn Tyr Asp Pro Glu Gly Asn Glu Ile Val
435 440 445
Gln His Lys Asn Trp Ser Glu Asn Asn Lys Ser Lys Leu Ala His Phe
450 455 460
Thr Ser Ser Ile Tyr Leu Pro Gly Asn Ala Arg Asn Ile Asn Val Tyr
465 470 475 480
Ala Lys Glu Cys Thr Gly Leu Ala Trp Glu Trp Trp Arg Thr Val Ile
485 490 495
Asp Asp Arg Asn Leu Pro Leu Val Lys Asn Arg Asn Ile Ser Ile Trp
500 505 510
Gly Thr Thr Leu Tyr Pro Lys Tyr Ser Asn Lys Val Asp Asn Pro Ile
515 520 525
Glu
<210> 57
<211> 441
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 57
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Lys Asp Ala Ser Ala Phe Asn Lys
20 25 30
Glu Asn Ser Ile Ser Ser Val Ala Pro Pro Ala Ser Pro Pro Ala Ser
35 40 45
Pro Lys Thr Pro Ile Glu Lys Lys His Ala Asp Glu Ile Asp Lys Tyr
50 55 60
Ile Gln Gly Leu Asp Tyr Asn Lys Asn Asn Val Leu Val Tyr His Gly
65 70 75 80
Asp Ala Val Thr Asn Val Pro Pro Arg Lys Gly Tyr Lys Asp Gly Asn
85 90 95
Glu Tyr Ile Val Val Glu Lys Lys Lys Lys Ser Ile Asn Gln Asn Asn
100 105 110
Ala Asp Ile Gln Val Val Asn Ala Ile Ser Ser Leu Thr Tyr Pro Gly
115 120 125
Ala Leu Val Lys Ala Asn Ser Glu Leu Val Glu Asn Gln Pro Asp Val
130 135 140
Leu Pro Val Lys Arg Asp Ser Leu Thr Leu Ser Ile Asp Leu Pro Gly
145 150 155 160
Met Thr Asn Gln Asp Asn Lys Ile Val Val Lys Asn Ala Thr Lys Ser
165 170 175
Asn Val Asn Asn Ala Val Asn Thr Leu Val Glu Arg Trp Asn Glu Lys
180 185 190
Tyr Ala Gln Ala Tyr Ser Asn Val Ser Ala Lys Ile Asp Tyr Asp Asp
195 200 205
Glu Met Ala Tyr Ser Glu Ser Gln Leu Ile Ala Lys Phe Gly Thr Ala
210 215 220
Phe Lys Ala Val Asn Asn Ser Leu Asn Val Asn Phe Gly Ala Ile Ser
225 230 235 240
Glu Gly Lys Met Gln Glu Glu Val Ile Ser Phe Lys Gln Ile Tyr Tyr
245 250 255
Asn Val Asn Val Asn Glu Pro Thr Arg Pro Ser Arg Phe Phe Gly Lys
260 265 270
Ala Val Thr Lys Glu Gln Leu Gln Ala Leu Gly Val Asn Ala Glu Asn
275 280 285
Pro Pro Ala Tyr Ile Ser Ser Val Ala Tyr Gly Arg Gln Val Tyr Leu
290 295 300
Lys Leu Ser Thr Asn Ser His Ser Thr Lys Val Lys Ala Ala Phe Asp
305 310 315 320
Ala Ala Val Ser Gly Lys Ser Val Ser Gly Asp Val Glu Leu Thr Asn
325 330 335
Ile Ile Lys Asn Ser Ser Phe Lys Ala Val Ile Tyr Gly Gly Ser Ala
340 345 350
Lys Asp Glu Val Gln Ile Ile Asp Gly Asn Leu Gly Asp Leu Arg Asp
355 360 365
Ile Leu Lys Lys Gly Ala Thr Phe Asn Arg Glu Thr Pro Gly Val Pro
370 375 380
Ile Ala Tyr Thr Thr Asn Phe Leu Lys Asp Asn Glu Leu Ala Val Ile
385 390 395 400
Lys Asn Asn Ser Glu Tyr Ile Glu Thr Thr Ser Lys Ala Tyr Thr Asp
405 410 415
Gly Lys Ile Asn Ile Asp His Ser Gly Gly Tyr Val Ala Gln Phe Asn
420 425 430
Ile Ser Trp Asp Glu Val Asn Tyr Asp
435 440
<210> 58
<211> 416
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 58
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Lys Asp Ala Ser Ala Phe Asn Lys
20 25 30
Glu Asn Ser Ile Ser Ser Val Ala Pro Pro Ala Ser Pro Pro Ala Ser
35 40 45
Pro Lys Thr Pro Ile Glu Lys Lys His Ala Asp Glu Ile Asp Lys Tyr
50 55 60
Ile Gln Gly Leu Asp Tyr Asn Lys Asn Asn Val Leu Val Tyr His Gly
65 70 75 80
Asp Ala Val Thr Asn Val Pro Pro Arg Lys Gly Tyr Lys Asp Gly Asn
85 90 95
Glu Tyr Ile Val Val Glu Lys Lys Lys Lys Ser Ile Asn Gln Asn Asn
100 105 110
Ala Asp Ile Gln Val Val Asn Ala Ile Ser Ser Leu Thr Tyr Pro Gly
115 120 125
Ala Leu Val Lys Ala Asn Ser Glu Leu Val Glu Asn Gln Pro Asp Val
130 135 140
Leu Pro Val Lys Arg Asp Ser Leu Thr Leu Ser Ile Asp Leu Pro Gly
145 150 155 160
Met Thr Asn Gln Asp Asn Lys Ile Val Val Lys Asn Ala Thr Lys Ser
165 170 175
Asn Val Asn Asn Ala Val Asn Thr Leu Val Glu Arg Trp Asn Glu Lys
180 185 190
Tyr Ala Gln Ala Tyr Ser Asn Val Ser Ala Lys Ile Asp Tyr Asp Asp
195 200 205
Glu Met Ala Tyr Ser Glu Ser Gln Leu Ile Ala Lys Phe Gly Thr Ala
210 215 220
Phe Lys Ala Val Asn Asn Ser Leu Asn Val Asn Phe Gly Ala Ile Ser
225 230 235 240
Glu Gly Lys Met Gln Glu Glu Val Ile Ser Phe Lys Gln Ile Tyr Tyr
245 250 255
Asn Val Asn Val Asn Glu Pro Thr Arg Pro Ser Arg Phe Phe Gly Lys
260 265 270
Ala Val Thr Lys Glu Gln Leu Gln Ala Leu Gly Val Asn Ala Glu Asn
275 280 285
Pro Pro Ala Tyr Ile Ser Ser Val Ala Tyr Gly Arg Gln Val Tyr Leu
290 295 300
Lys Leu Ser Thr Asn Ser His Ser Thr Lys Val Lys Ala Ala Phe Asp
305 310 315 320
Ala Ala Val Ser Gly Lys Ser Val Ser Gly Asp Val Glu Leu Thr Asn
325 330 335
Ile Ile Lys Asn Ser Ser Phe Lys Ala Val Ile Tyr Gly Gly Ser Ala
340 345 350
Lys Asp Glu Val Gln Ile Ile Asp Gly Asn Leu Gly Asp Leu Arg Asp
355 360 365
Ile Leu Lys Lys Gly Ala Thr Phe Asn Arg Glu Thr Pro Gly Val Pro
370 375 380
Ile Ala Tyr Thr Thr Asn Phe Leu Lys Asp Asn Glu Leu Ala Val Ile
385 390 395 400
Lys Asn Asn Ser Glu Tyr Ile Glu Thr Thr Ser Lys Ala Tyr Thr Asp
405 410 415
<210> 59
<211> 441
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 59
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Lys Asp Ala Ser Ala Phe Asn Lys
20 25 30
Glu Asn Ser Ile Ser Ser Met Ala Pro Pro Ala Ser Pro Pro Ala Ser
35 40 45
Pro Lys Thr Pro Ile Glu Lys Lys His Ala Asp Glu Ile Asp Lys Tyr
50 55 60
Ile Gln Gly Leu Asp Tyr Asn Lys Asn Asn Val Leu Val Tyr His Gly
65 70 75 80
Asp Ala Val Thr Asn Val Pro Pro Arg Lys Gly Tyr Lys Asp Gly Asn
85 90 95
Glu Tyr Ile Val Val Glu Lys Lys Lys Lys Ser Ile Asn Gln Asn Asn
100 105 110
Ala Asp Ile Gln Val Val Asn Ala Ile Ser Ser Leu Thr Tyr Pro Gly
115 120 125
Ala Leu Val Lys Ala Asn Ser Glu Leu Val Glu Asn Gln Pro Asp Val
130 135 140
Leu Pro Val Lys Arg Asp Ser Leu Thr Leu Ser Ile Asp Leu Pro Gly
145 150 155 160
Met Thr Asn Gln Asp Asn Lys Ile Val Val Lys Asn Ala Thr Lys Ser
165 170 175
Asn Val Asn Asn Ala Val Asn Thr Leu Val Glu Arg Trp Asn Glu Lys
180 185 190
Tyr Ala Gln Ala Tyr Pro Asn Val Ser Ala Lys Ile Asp Tyr Asp Asp
195 200 205
Glu Met Ala Tyr Ser Glu Ser Gln Leu Ile Ala Lys Phe Gly Thr Ala
210 215 220
Phe Lys Ala Val Asn Asn Ser Leu Asn Val Asn Phe Gly Ala Ile Ser
225 230 235 240
Glu Gly Lys Met Gln Glu Glu Val Ile Ser Phe Lys Gln Ile Tyr Tyr
245 250 255
Asn Val Asn Val Asn Glu Pro Thr Arg Pro Ser Arg Phe Phe Gly Lys
260 265 270
Ala Val Thr Lys Glu Gln Leu Gln Ala Leu Gly Val Asn Ala Glu Asn
275 280 285
Pro Pro Ala Tyr Ile Ser Ser Val Ala Tyr Gly Arg Gln Val Tyr Leu
290 295 300
Lys Leu Ser Thr Asn Ser His Ser Thr Lys Val Lys Ala Ala Phe Asp
305 310 315 320
Ala Ala Val Ser Gly Lys Ser Val Ser Gly Asp Val Glu Leu Thr Asn
325 330 335
Ile Ile Lys Asn Ser Ser Phe Lys Ala Val Ile Tyr Gly Gly Ser Ala
340 345 350
Lys Asp Glu Val Gln Ile Ile Asp Gly Asn Leu Gly Asp Leu Arg Asp
355 360 365
Ile Leu Lys Lys Gly Ala Thr Phe Asn Arg Glu Thr Pro Gly Val Pro
370 375 380
Ile Ala Tyr Thr Thr Asn Phe Leu Lys Asp Asn Glu Leu Ala Val Ile
385 390 395 400
Lys Asn Asn Ser Glu Tyr Ile Glu Thr Thr Ser Lys Ala Tyr Thr Asp
405 410 415
Gly Lys Ile Asn Ile Asp His Ser Gly Gly Tyr Val Ala Gln Phe Asn
420 425 430
Ile Ser Trp Asp Glu Val Asn Tyr Asp
435 440
<210> 60
<211> 1323
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 60
atgaaaaaaa taatgctagt ttttattaca cttatattag ttagtctacc aattgcgcaa 60
caaactgaag caaaggatgc atctgcattc aataaagaaa attcaatttc atccatggca 120
ccaccagcat ctccgcctgc aagtcctaag acgccaatcg aaaagaaaca cgcggatgaa 180
atcgataagt atatacaagg attggattac aataaaaaca atgtattagt ataccacgga 240
gatgcagtga caaatgtgcc gccaagaaaa ggttacaaag atggaaatga atatattgtt 300
gtggagaaaa agaagaaatc catcaatcaa aataatgcag acattcaagt tgtgaatgca 360
atttcgagcc taacctatcc aggtgctctc gtaaaagcga attcggaatt agtagaaaat 420
caaccagatg ttctccctgt aaaacgtgat tcattaacac tcagcattga tttgccaggt 480
atgactaatc aagacaataa aatagttgta aaaaatgcca ctaaatcaaa cgttaacaac 540
gcagtaaata cattagtgga aagatggaat gaaaaatatg ctcaagctta tccaaatgta 600
agtgcaaaaa ttgattatga tgacgaaatg gcttacagtg aatcacaatt aattgcgaaa 660
tttggtacag catttaaagc tgtaaataat agcttgaatg taaacttcgg cgcaatcagt 720
gaagggaaaa tgcaagaaga agtcattagt tttaaacaaa tttactataa cgtgaatgtt 780
aatgaaccta caagaccttc cagatttttc ggcaaagctg ttactaaaga gcagttgcaa 840
gcgcttggag tgaatgcaga aaatcctcct gcatatatct caagtgtggc gtatggccgt 900
caagtttatt tgaaattatc aactaattcc catagtacta aagtaaaagc tgcttttgat 960
gctgccgtaa gcggaaaatc tgtctcaggt gatgtagaac taacaaatat catcaaaaat 1020
tcttccttca aagccgtaat ttacggaggt tccgcaaaag atgaagttca aatcatcgac 1080
ggcaacctcg gagacttacg cgatattttg aaaaaaggcg ctacttttaa tcgagaaaca 1140
ccaggagttc ccattgctta tacaacaaac ttcctaaaag acaatgaatt agctgttatt 1200
aaaaacaact cagaatatat tgaaacaact tcaaaagctt atacagatgg aaaaattaac 1260
atcgatcact ctggaggata cgttgctcaa ttcaacattt cttgggatga agtaaattat 1320
gat 1323
<210> 61
<211> 633
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 61
Met Arg Ala Met Met Val Val Phe Ile Thr Ala Asn Cys Ile Thr Ile
1 5 10 15
Asn Pro Asp Ile Ile Phe Ala Ala Thr Asp Ser Glu Asp Ser Ser Leu
20 25 30
Asn Thr Asp Glu Trp Glu Glu Glu Lys Thr Glu Glu Gln Pro Ser Glu
35 40 45
Val Asn Thr Gly Pro Arg Tyr Glu Thr Ala Arg Glu Val Ser Ser Arg
50 55 60
Asp Ile Glu Glu Leu Glu Lys Ser Asn Lys Val Lys Asn Thr Asn Lys
65 70 75 80
Ala Asp Leu Ile Ala Met Leu Lys Ala Lys Ala Glu Lys Gly Pro Asn
85 90 95
Asn Asn Asn Asn Asn Gly Glu Gln Thr Gly Asn Val Ala Ile Asn Glu
100 105 110
Glu Ala Ser Gly Val Asp Arg Pro Thr Leu Gln Val Glu Arg Arg His
115 120 125
Pro Gly Leu Ser Ser Asp Ser Ala Ala Glu Ile Lys Lys Arg Arg Lys
130 135 140
Ala Ile Ala Ser Ser Asp Ser Glu Leu Glu Ser Leu Thr Tyr Pro Asp
145 150 155 160
Lys Pro Thr Lys Ala Asn Lys Arg Lys Val Ala Lys Glu Ser Val Val
165 170 175
Asp Ala Ser Glu Ser Asp Leu Asp Ser Ser Met Gln Ser Ala Asp Glu
180 185 190
Ser Thr Pro Gln Pro Leu Lys Ala Asn Gln Lys Pro Phe Phe Pro Lys
195 200 205
Val Phe Lys Lys Ile Lys Asp Ala Gly Lys Trp Val Arg Asp Lys Ile
210 215 220
Asp Glu Asn Pro Glu Val Lys Lys Ala Ile Val Asp Lys Ser Ala Gly
225 230 235 240
Leu Ile Asp Gln Leu Leu Thr Lys Lys Lys Ser Glu Glu Val Asn Ala
245 250 255
Ser Asp Phe Pro Pro Pro Pro Thr Asp Glu Glu Leu Arg Leu Ala Leu
260 265 270
Pro Glu Thr Pro Met Leu Leu Gly Phe Asn Ala Pro Thr Pro Ser Glu
275 280 285
Pro Ser Ser Phe Glu Phe Pro Pro Pro Pro Thr Asp Glu Glu Leu Arg
290 295 300
Leu Ala Leu Pro Glu Thr Pro Met Leu Leu Gly Phe Asn Ala Pro Ala
305 310 315 320
Thr Ser Glu Pro Ser Ser Phe Glu Phe Pro Pro Pro Pro Thr Glu Asp
325 330 335
Glu Leu Glu Ile Met Arg Glu Thr Ala Pro Ser Leu Asp Ser Ser Phe
340 345 350
Thr Ser Gly Asp Leu Ala Ser Leu Arg Ser Ala Ile Asn Arg His Ser
355 360 365
Glu Asn Phe Ser Asp Phe Pro Leu Ile Pro Thr Glu Glu Glu Leu Asn
370 375 380
Gly Arg Gly Gly Arg Pro Thr Ser Glu Glu Phe Ser Ser Leu Asn Ser
385 390 395 400
Gly Asp Phe Thr Asp Asp Glu Asn Ser Glu Thr Thr Glu Glu Glu Ile
405 410 415
Asp Arg Leu Ala Asp Leu Arg Asp Arg Gly Thr Gly Lys His Ser Arg
420 425 430
Asn Ala Gly Phe Leu Pro Leu Asn Pro Phe Ile Ser Ser Pro Val Pro
435 440 445
Ser Leu Thr Pro Lys Val Pro Lys Ile Ser Ala Pro Ala Leu Ile Ser
450 455 460
Asp Ile Thr Lys Lys Ala Pro Phe Lys Asn Pro Ser Gln Pro Leu Asn
465 470 475 480
Val Phe Asn Lys Lys Thr Thr Thr Lys Thr Val Thr Lys Lys Pro Thr
485 490 495
Pro Val Lys Thr Ala Pro Lys Leu Ala Glu Leu Pro Ala Thr Lys Pro
500 505 510
Gln Glu Thr Val Leu Arg Glu Asn Lys Thr Pro Phe Ile Glu Lys Gln
515 520 525
Ala Glu Thr Asn Lys Gln Ser Ile Asn Met Pro Ser Leu Pro Val Ile
530 535 540
Gln Lys Glu Ala Thr Glu Ser Asp Lys Glu Glu Met Lys Pro Gln Thr
545 550 555 560
Glu Glu Lys Met Val Glu Glu Ser Glu Ser Ala Asn Asn Ala Asn Gly
565 570 575
Lys Asn Arg Ser Ala Gly Ile Glu Glu Gly Lys Leu Ile Ala Lys Ser
580 585 590
Ala Glu Asp Glu Lys Ala Lys Glu Glu Pro Gly Asn His Thr Thr Leu
595 600 605
Ile Leu Ala Met Leu Ala Ile Gly Val Phe Ser Leu Gly Ala Phe Ile
610 615 620
Lys Ile Ile Gln Leu Arg Lys Asn Asn
625 630
<210> 62
<211> 639
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 62
Met Gly Leu Asn Arg Phe Met Arg Ala Met Met Val Val Phe Ile Thr
1 5 10 15
Ala Asn Cys Ile Thr Ile Asn Pro Asp Ile Ile Phe Ala Ala Thr Asp
20 25 30
Ser Glu Asp Ser Ser Leu Asn Thr Asp Glu Trp Glu Glu Glu Lys Thr
35 40 45
Glu Glu Gln Pro Ser Glu Val Asn Thr Gly Pro Arg Tyr Glu Thr Ala
50 55 60
Arg Glu Val Ser Ser Arg Asp Ile Glu Glu Leu Glu Lys Ser Asn Lys
65 70 75 80
Val Lys Asn Thr Asn Lys Ala Asp Leu Ile Ala Met Leu Lys Ala Lys
85 90 95
Ala Glu Lys Gly Pro Asn Asn Asn Asn Asn Asn Gly Glu Gln Thr Gly
100 105 110
Asn Val Ala Ile Asn Glu Glu Ala Ser Gly Val Asp Arg Pro Thr Leu
115 120 125
Gln Val Glu Arg Arg His Pro Gly Leu Ser Ser Asp Ser Ala Ala Glu
130 135 140
Ile Lys Lys Arg Arg Lys Ala Ile Ala Ser Ser Asp Ser Glu Leu Glu
145 150 155 160
Ser Leu Thr Tyr Pro Asp Lys Pro Thr Lys Ala Asn Lys Arg Lys Val
165 170 175
Ala Lys Glu Ser Val Val Asp Ala Ser Glu Ser Asp Leu Asp Ser Ser
180 185 190
Met Gln Ser Ala Asp Glu Ser Thr Pro Gln Pro Leu Lys Ala Asn Gln
195 200 205
Lys Pro Phe Phe Pro Lys Val Phe Lys Lys Ile Lys Asp Ala Gly Lys
210 215 220
Trp Val Arg Asp Lys Ile Asp Glu Asn Pro Glu Val Lys Lys Ala Ile
225 230 235 240
Val Asp Lys Ser Ala Gly Leu Ile Asp Gln Leu Leu Thr Lys Lys Lys
245 250 255
Ser Glu Glu Val Asn Ala Ser Asp Phe Pro Pro Pro Pro Thr Asp Glu
260 265 270
Glu Leu Arg Leu Ala Leu Pro Glu Thr Pro Met Leu Leu Gly Phe Asn
275 280 285
Ala Pro Thr Pro Ser Glu Pro Ser Ser Phe Glu Phe Pro Pro Pro Pro
290 295 300
Thr Asp Glu Glu Leu Arg Leu Ala Leu Pro Glu Thr Pro Met Leu Leu
305 310 315 320
Gly Phe Asn Ala Pro Ala Thr Ser Glu Pro Ser Ser Phe Glu Phe Pro
325 330 335
Pro Pro Pro Thr Glu Asp Glu Leu Glu Ile Met Arg Glu Thr Ala Pro
340 345 350
Ser Leu Asp Ser Ser Phe Thr Ser Gly Asp Leu Ala Ser Leu Arg Ser
355 360 365
Ala Ile Asn Arg His Ser Glu Asn Phe Ser Asp Phe Pro Leu Ile Pro
370 375 380
Thr Glu Glu Glu Leu Asn Gly Arg Gly Gly Arg Pro Thr Ser Glu Glu
385 390 395 400
Phe Ser Ser Leu Asn Ser Gly Asp Phe Thr Asp Asp Glu Asn Ser Glu
405 410 415
Thr Thr Glu Glu Glu Ile Asp Arg Leu Ala Asp Leu Arg Asp Arg Gly
420 425 430
Thr Gly Lys His Ser Arg Asn Ala Gly Phe Leu Pro Leu Asn Pro Phe
435 440 445
Ile Ser Ser Pro Val Pro Ser Leu Thr Pro Lys Val Pro Lys Ile Ser
450 455 460
Ala Pro Ala Leu Ile Ser Asp Ile Thr Lys Lys Ala Pro Phe Lys Asn
465 470 475 480
Pro Ser Gln Pro Leu Asn Val Phe Asn Lys Lys Thr Thr Thr Lys Thr
485 490 495
Val Thr Lys Lys Pro Thr Pro Val Lys Thr Ala Pro Lys Leu Ala Glu
500 505 510
Leu Pro Ala Thr Lys Pro Gln Glu Thr Val Leu Arg Glu Asn Lys Thr
515 520 525
Pro Phe Ile Glu Lys Gln Ala Glu Thr Asn Lys Gln Ser Ile Asn Met
530 535 540
Pro Ser Leu Pro Val Ile Gln Lys Glu Ala Thr Glu Ser Asp Lys Glu
545 550 555 560
Glu Met Lys Pro Gln Thr Glu Glu Lys Met Val Glu Glu Ser Glu Ser
565 570 575
Ala Asn Asn Ala Asn Gly Lys Asn Arg Ser Ala Gly Ile Glu Glu Gly
580 585 590
Lys Leu Ile Ala Lys Ser Ala Glu Asp Glu Lys Ala Lys Glu Glu Pro
595 600 605
Gly Asn His Thr Thr Leu Ile Leu Ala Met Leu Ala Ile Gly Val Phe
610 615 620
Ser Leu Gly Ala Phe Ile Lys Ile Ile Gln Leu Arg Lys Asn Asn
625 630 635
<210> 63
<211> 93
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 63
Ala Thr Asp Ser Glu Asp Ser Ser Leu Asn Thr Asp Glu Trp Glu Glu
1 5 10 15
Glu Lys Thr Glu Glu Gln Pro Ser Glu Val Asn Thr Gly Pro Arg Tyr
20 25 30
Glu Thr Ala Arg Glu Val Ser Ser Arg Asp Ile Glu Glu Leu Glu Lys
35 40 45
Ser Asn Lys Val Lys Asn Thr Asn Lys Ala Asp Leu Ile Ala Met Leu
50 55 60
Lys Ala Lys Ala Glu Lys Gly Pro Asn Asn Asn Asn Asn Asn Gly Glu
65 70 75 80
Gln Thr Gly Asn Val Ala Ile Asn Glu Glu Ala Ser Gly
85 90
<210> 64
<211> 200
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 64
Ala Thr Asp Ser Glu Asp Ser Ser Leu Asn Thr Asp Glu Trp Glu Glu
1 5 10 15
Glu Lys Thr Glu Glu Gln Pro Ser Glu Val Asn Thr Gly Pro Arg Tyr
20 25 30
Glu Thr Ala Arg Glu Val Ser Ser Arg Asp Ile Glu Glu Leu Glu Lys
35 40 45
Ser Asn Lys Val Lys Asn Thr Asn Lys Ala Asp Leu Ile Ala Met Leu
50 55 60
Lys Ala Lys Ala Glu Lys Gly Pro Asn Asn Asn Asn Asn Asn Gly Glu
65 70 75 80
Gln Thr Gly Asn Val Ala Ile Asn Glu Glu Ala Ser Gly Val Asp Arg
85 90 95
Pro Thr Leu Gln Val Glu Arg Arg His Pro Gly Leu Ser Ser Asp Ser
100 105 110
Ala Ala Glu Ile Lys Lys Arg Arg Lys Ala Ile Ala Ser Ser Asp Ser
115 120 125
Glu Leu Glu Ser Leu Thr Tyr Pro Asp Lys Pro Thr Lys Ala Asn Lys
130 135 140
Arg Lys Val Ala Lys Glu Ser Val Val Asp Ala Ser Glu Ser Asp Leu
145 150 155 160
Asp Ser Ser Met Gln Ser Ala Asp Glu Ser Thr Pro Gln Pro Leu Lys
165 170 175
Ala Asn Gln Lys Pro Phe Phe Pro Lys Val Phe Lys Lys Ile Lys Asp
180 185 190
Ala Gly Lys Trp Val Arg Asp Lys
195 200
<210> 65
<211> 303
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 65
Ala Thr Asp Ser Glu Asp Ser Ser Leu Asn Thr Asp Glu Trp Glu Glu
1 5 10 15
Glu Lys Thr Glu Glu Gln Pro Ser Glu Val Asn Thr Gly Pro Arg Tyr
20 25 30
Glu Thr Ala Arg Glu Val Ser Ser Arg Asp Ile Glu Glu Leu Glu Lys
35 40 45
Ser Asn Lys Val Lys Asn Thr Asn Lys Ala Asp Leu Ile Ala Met Leu
50 55 60
Lys Ala Lys Ala Glu Lys Gly Pro Asn Asn Asn Asn Asn Asn Gly Glu
65 70 75 80
Gln Thr Gly Asn Val Ala Ile Asn Glu Glu Ala Ser Gly Val Asp Arg
85 90 95
Pro Thr Leu Gln Val Glu Arg Arg His Pro Gly Leu Ser Ser Asp Ser
100 105 110
Ala Ala Glu Ile Lys Lys Arg Arg Lys Ala Ile Ala Ser Ser Asp Ser
115 120 125
Glu Leu Glu Ser Leu Thr Tyr Pro Asp Lys Pro Thr Lys Ala Asn Lys
130 135 140
Arg Lys Val Ala Lys Glu Ser Val Val Asp Ala Ser Glu Ser Asp Leu
145 150 155 160
Asp Ser Ser Met Gln Ser Ala Asp Glu Ser Thr Pro Gln Pro Leu Lys
165 170 175
Ala Asn Gln Lys Pro Phe Phe Pro Lys Val Phe Lys Lys Ile Lys Asp
180 185 190
Ala Gly Lys Trp Val Arg Asp Lys Ile Asp Glu Asn Pro Glu Val Lys
195 200 205
Lys Ala Ile Val Asp Lys Ser Ala Gly Leu Ile Asp Gln Leu Leu Thr
210 215 220
Lys Lys Lys Ser Glu Glu Val Asn Ala Ser Asp Phe Pro Pro Pro Pro
225 230 235 240
Thr Asp Glu Glu Leu Arg Leu Ala Leu Pro Glu Thr Pro Met Leu Leu
245 250 255
Gly Phe Asn Ala Pro Thr Pro Ser Glu Pro Ser Ser Phe Glu Phe Pro
260 265 270
Pro Pro Pro Thr Asp Glu Glu Leu Arg Leu Ala Leu Pro Glu Thr Pro
275 280 285
Met Leu Leu Gly Phe Asn Ala Pro Ala Thr Ser Glu Pro Ser Ser
290 295 300
<210> 66
<211> 370
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 66
Ala Thr Asp Ser Glu Asp Ser Ser Leu Asn Thr Asp Glu Trp Glu Glu
1 5 10 15
Glu Lys Thr Glu Glu Gln Pro Ser Glu Val Asn Thr Gly Pro Arg Tyr
20 25 30
Glu Thr Ala Arg Glu Val Ser Ser Arg Asp Ile Glu Glu Leu Glu Lys
35 40 45
Ser Asn Lys Val Lys Asn Thr Asn Lys Ala Asp Leu Ile Ala Met Leu
50 55 60
Lys Ala Lys Ala Glu Lys Gly Pro Asn Asn Asn Asn Asn Asn Gly Glu
65 70 75 80
Gln Thr Gly Asn Val Ala Ile Asn Glu Glu Ala Ser Gly Val Asp Arg
85 90 95
Pro Thr Leu Gln Val Glu Arg Arg His Pro Gly Leu Ser Ser Asp Ser
100 105 110
Ala Ala Glu Ile Lys Lys Arg Arg Lys Ala Ile Ala Ser Ser Asp Ser
115 120 125
Glu Leu Glu Ser Leu Thr Tyr Pro Asp Lys Pro Thr Lys Ala Asn Lys
130 135 140
Arg Lys Val Ala Lys Glu Ser Val Val Asp Ala Ser Glu Ser Asp Leu
145 150 155 160
Asp Ser Ser Met Gln Ser Ala Asp Glu Ser Thr Pro Gln Pro Leu Lys
165 170 175
Ala Asn Gln Lys Pro Phe Phe Pro Lys Val Phe Lys Lys Ile Lys Asp
180 185 190
Ala Gly Lys Trp Val Arg Asp Lys Ile Asp Glu Asn Pro Glu Val Lys
195 200 205
Lys Ala Ile Val Asp Lys Ser Ala Gly Leu Ile Asp Gln Leu Leu Thr
210 215 220
Lys Lys Lys Ser Glu Glu Val Asn Ala Ser Asp Phe Pro Pro Pro Pro
225 230 235 240
Thr Asp Glu Glu Leu Arg Leu Ala Leu Pro Glu Thr Pro Met Leu Leu
245 250 255
Gly Phe Asn Ala Pro Thr Pro Ser Glu Pro Ser Ser Phe Glu Phe Pro
260 265 270
Pro Pro Pro Thr Asp Glu Glu Leu Arg Leu Ala Leu Pro Glu Thr Pro
275 280 285
Met Leu Leu Gly Phe Asn Ala Pro Ala Thr Ser Glu Pro Ser Ser Phe
290 295 300
Glu Phe Pro Pro Pro Pro Thr Glu Asp Glu Leu Glu Ile Met Arg Glu
305 310 315 320
Thr Ala Pro Ser Leu Asp Ser Ser Phe Thr Ser Gly Asp Leu Ala Ser
325 330 335
Leu Arg Ser Ala Ile Asn Arg His Ser Glu Asn Phe Ser Asp Phe Pro
340 345 350
Leu Ile Pro Thr Glu Glu Glu Leu Asn Gly Arg Gly Gly Arg Pro Thr
355 360 365
Ser Glu
370
<210> 67
<211> 390
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 67
Met Arg Ala Met Met Val Val Phe Ile Thr Ala Asn Cys Ile Thr Ile
1 5 10 15
Asn Pro Asp Ile Ile Phe Ala Ala Thr Asp Ser Glu Asp Ser Ser Leu
20 25 30
Asn Thr Asp Glu Trp Glu Glu Glu Lys Thr Glu Glu Gln Pro Ser Glu
35 40 45
Val Asn Thr Gly Pro Arg Tyr Glu Thr Ala Arg Glu Val Ser Ser Arg
50 55 60
Asp Ile Lys Glu Leu Glu Lys Ser Asn Lys Val Arg Asn Thr Asn Lys
65 70 75 80
Ala Asp Leu Ile Ala Met Leu Lys Glu Lys Ala Glu Lys Gly Pro Asn
85 90 95
Ile Asn Asn Asn Asn Ser Glu Gln Thr Glu Asn Ala Ala Ile Asn Glu
100 105 110
Glu Ala Ser Gly Ala Asp Arg Pro Ala Ile Gln Val Glu Arg Arg His
115 120 125
Pro Gly Leu Pro Ser Asp Ser Ala Ala Glu Ile Lys Lys Arg Arg Lys
130 135 140
Ala Ile Ala Ser Ser Asp Ser Glu Leu Glu Ser Leu Thr Tyr Pro Asp
145 150 155 160
Lys Pro Thr Lys Val Asn Lys Lys Lys Val Ala Lys Glu Ser Val Ala
165 170 175
Asp Ala Ser Glu Ser Asp Leu Asp Ser Ser Met Gln Ser Ala Asp Glu
180 185 190
Ser Ser Pro Gln Pro Leu Lys Ala Asn Gln Gln Pro Phe Phe Pro Lys
195 200 205
Val Phe Lys Lys Ile Lys Asp Ala Gly Lys Trp Val Arg Asp Lys Ile
210 215 220
Asp Glu Asn Pro Glu Val Lys Lys Ala Ile Val Asp Lys Ser Ala Gly
225 230 235 240
Leu Ile Asp Gln Leu Leu Thr Lys Lys Lys Ser Glu Glu Val Asn Ala
245 250 255
Ser Asp Phe Pro Pro Pro Pro Thr Asp Glu Glu Leu Arg Leu Ala Leu
260 265 270
Pro Glu Thr Pro Met Leu Leu Gly Phe Asn Ala Pro Ala Thr Ser Glu
275 280 285
Pro Ser Ser Phe Glu Phe Pro Pro Pro Pro Thr Asp Glu Glu Leu Arg
290 295 300
Leu Ala Leu Pro Glu Thr Pro Met Leu Leu Gly Phe Asn Ala Pro Ala
305 310 315 320
Thr Ser Glu Pro Ser Ser Phe Glu Phe Pro Pro Pro Pro Thr Glu Asp
325 330 335
Glu Leu Glu Ile Ile Arg Glu Thr Ala Ser Ser Leu Asp Ser Ser Phe
340 345 350
Thr Arg Gly Asp Leu Ala Ser Leu Arg Asn Ala Ile Asn Arg His Ser
355 360 365
Gln Asn Phe Ser Asp Phe Pro Pro Ile Pro Thr Glu Glu Glu Leu Asn
370 375 380
Gly Arg Gly Gly Arg Pro
385 390
<210> 68
<211> 1170
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 68
atgcgtgcga tgatggtggt tttcattact gccaattgca ttacgattaa ccccgacata 60
atatttgcag cgacagatag cgaagattct agtctaaaca cagatgaatg ggaagaagaa 120
aaaacagaag agcaaccaag cgaggtaaat acgggaccaa gatacgaaac tgcacgtgaa 180
gtaagttcac gtgatattaa agaactagaa aaatcgaata aagtgagaaa tacgaacaaa 240
gcagacctaa tagcaatgtt gaaagaaaaa gcagaaaaag gtccaaatat caataataac 300
aacagtgaac aaactgagaa tgcggctata aatgaagagg cttcaggagc cgaccgacca 360
gctatacaag tggagcgtcg tcatccagga ttgccatcgg atagcgcagc ggaaattaaa 420
aaaagaagga aagccatagc atcatcggat agtgagcttg aaagccttac ttatccggat 480
aaaccaacaa aagtaaataa gaaaaaagtg gcgaaagagt cagttgcgga tgcttctgaa 540
agtgacttag attctagcat gcagtcagca gatgagtctt caccacaacc tttaaaagca 600
aaccaacaac catttttccc taaagtattt aaaaaaataa aagatgcggg gaaatgggta 660
cgtgataaaa tcgacgaaaa tcctgaagta aagaaagcga ttgttgataa aagtgcaggg 720
ttaattgacc aattattaac caaaaagaaa agtgaagagg taaatgcttc ggacttcccg 780
ccaccaccta cggatgaaga gttaagactt gctttgccag agacaccaat gcttcttggt 840
tttaatgctc ctgctacatc agaaccgagc tcattcgaat ttccaccacc acctacggat 900
gaagagttaa gacttgcttt gccagagacg ccaatgcttc ttggttttaa tgctcctgct 960
acatcggaac cgagctcgtt cgaatttcca ccgcctccaa cagaagatga actagaaatc 1020
atccgggaaa cagcatcctc gctagattct agttttacaa gaggggattt agctagtttg 1080
agaaatgcta ttaatcgcca tagtcaaaat ttctctgatt tcccaccaat cccaacagaa 1140
gaagagttga acgggagagg cggtagacca 1170
<210> 69
<211> 100
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 69
Met Gly Leu Asn Arg Phe Met Arg Ala Met Met Val Val Phe Ile Thr
1 5 10 15
Ala Asn Cys Ile Thr Ile Asn Pro Asp Ile Ile Phe Ala Ala Thr Asp
20 25 30
Ser Glu Asp Ser Ser Leu Asn Thr Asp Glu Trp Glu Glu Glu Lys Thr
35 40 45
Glu Glu Gln Pro Ser Glu Val Asn Thr Gly Pro Arg Tyr Glu Thr Ala
50 55 60
Arg Glu Val Ser Ser Arg Asp Ile Lys Glu Leu Glu Lys Ser Asn Lys
65 70 75 80
Val Arg Asn Thr Asn Lys Ala Asp Leu Ile Ala Met Leu Lys Glu Lys
85 90 95
Ala Glu Lys Gly
100
<210> 70
<211> 390
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 70
Met Arg Ala Met Met Val Val Phe Ile Thr Ala Asn Cys Ile Thr Ile
1 5 10 15
Asn Pro Asp Ile Ile Phe Ala Ala Thr Asp Ser Glu Asp Ser Ser Leu
20 25 30
Asn Thr Asp Glu Trp Glu Glu Glu Lys Thr Glu Glu Gln Pro Ser Glu
35 40 45
Val Asn Thr Gly Pro Arg Tyr Glu Thr Ala Arg Glu Val Ser Ser Arg
50 55 60
Asp Ile Glu Glu Leu Glu Lys Ser Asn Lys Val Lys Asn Thr Asn Lys
65 70 75 80
Ala Asp Leu Ile Ala Met Leu Lys Ala Lys Ala Glu Lys Gly Pro Asn
85 90 95
Asn Asn Asn Asn Asn Gly Glu Gln Thr Gly Asn Val Ala Ile Asn Glu
100 105 110
Glu Ala Ser Gly Val Asp Arg Pro Thr Leu Gln Val Glu Arg Arg His
115 120 125
Pro Gly Leu Ser Ser Asp Ser Ala Ala Glu Ile Lys Lys Arg Arg Lys
130 135 140
Ala Ile Ala Ser Ser Asp Ser Glu Leu Glu Ser Leu Thr Tyr Pro Asp
145 150 155 160
Lys Pro Thr Lys Ala Asn Lys Arg Lys Val Ala Lys Glu Ser Val Val
165 170 175
Asp Ala Ser Glu Ser Asp Leu Asp Ser Ser Met Gln Ser Ala Asp Glu
180 185 190
Ser Thr Pro Gln Pro Leu Lys Ala Asn Gln Lys Pro Phe Phe Pro Lys
195 200 205
Val Phe Lys Lys Ile Lys Asp Ala Gly Lys Trp Val Arg Asp Lys Ile
210 215 220
Asp Glu Asn Pro Glu Val Lys Lys Ala Ile Val Asp Lys Ser Ala Gly
225 230 235 240
Leu Ile Asp Gln Leu Leu Thr Lys Lys Lys Ser Glu Glu Val Asn Ala
245 250 255
Ser Asp Phe Pro Pro Pro Pro Thr Asp Glu Glu Leu Arg Leu Ala Leu
260 265 270
Pro Glu Thr Pro Met Leu Leu Gly Phe Asn Ala Pro Thr Pro Ser Glu
275 280 285
Pro Ser Ser Phe Glu Phe Pro Pro Pro Pro Thr Asp Glu Glu Leu Arg
290 295 300
Leu Ala Leu Pro Glu Thr Pro Met Leu Leu Gly Phe Asn Ala Pro Ala
305 310 315 320
Thr Ser Glu Pro Ser Ser Phe Glu Phe Pro Pro Pro Pro Thr Glu Asp
325 330 335
Glu Leu Glu Ile Met Arg Glu Thr Ala Pro Ser Leu Asp Ser Ser Phe
340 345 350
Thr Ser Gly Asp Leu Ala Ser Leu Arg Ser Ala Ile Asn Arg His Ser
355 360 365
Glu Asn Phe Ser Asp Phe Pro Leu Ile Pro Thr Glu Glu Glu Leu Asn
370 375 380
Gly Arg Gly Gly Arg Pro
385 390
<210> 71
<211> 1170
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 71
atgcgtgcga tgatggtagt tttcattact gccaactgca ttacgattaa ccccgacata 60
atatttgcag cgacagatag cgaagattcc agtctaaaca cagatgaatg ggaagaagaa 120
aaaacagaag agcagccaag cgaggtaaat acgggaccaa gatacgaaac tgcacgtgaa 180
gtaagttcac gtgatattga ggaactagaa aaatcgaata aagtgaaaaa tacgaacaaa 240
gcagacctaa tagcaatgtt gaaagcaaaa gcagagaaag gtccgaataa caataataac 300
aacggtgagc aaacaggaaa tgtggctata aatgaagagg cttcaggagt cgaccgacca 360
actctgcaag tggagcgtcg tcatccaggt ctgtcatcgg atagcgcagc ggaaattaaa 420
aaaagaagaa aagccatagc gtcgtcggat agtgagcttg aaagccttac ttatccagat 480
aaaccaacaa aagcaaataa gagaaaagtg gcgaaagagt cagttgtgga tgcttctgaa 540
agtgacttag attctagcat gcagtcagca gacgagtcta caccacaacc tttaaaagca 600
aatcaaaaac catttttccc taaagtattt aaaaaaataa aagatgcggg gaaatgggta 660
cgtgataaaa tcgacgaaaa tcctgaagta aagaaagcga ttgttgataa aagtgcaggg 720
ttaattgacc aattattaac caaaaagaaa agtgaagagg taaatgcttc ggacttcccg 780
ccaccaccta cggatgaaga gttaagactt gctttgccag agacaccgat gcttctcggt 840
tttaatgctc ctactccatc ggaaccgagc tcattcgaat ttccgccgcc acctacggat 900
gaagagttaa gacttgcttt gccagagacg ccaatgcttc ttggttttaa tgctcctgct 960
acatcggaac cgagctcatt cgaatttcca ccgcctccaa cagaagatga actagaaatt 1020
atgcgggaaa cagcaccttc gctagattct agttttacaa gcggggattt agctagtttg 1080
agaagtgcta ttaatcgcca tagcgaaaat ttctctgatt tcccactaat cccaacagaa 1140
gaagagttga acgggagagg cggtagacca 1170
<210> 72
<211> 226
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 72
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Ser Arg Ala Thr Asp Ser Glu Asp
20 25 30
Ser Ser Leu Asn Thr Asp Glu Trp Glu Glu Glu Lys Thr Glu Glu Gln
35 40 45
Pro Ser Glu Val Asn Thr Gly Pro Arg Tyr Glu Thr Ala Arg Glu Val
50 55 60
Ser Ser Arg Asp Ile Glu Glu Leu Glu Lys Ser Asn Lys Val Lys Asn
65 70 75 80
Thr Asn Lys Ala Asp Leu Ile Ala Met Leu Lys Ala Lys Ala Glu Lys
85 90 95
Gly Pro Asn Asn Asn Asn Asn Asn Gly Glu Gln Thr Gly Asn Val Ala
100 105 110
Ile Asn Glu Glu Ala Ser Gly Val Asp Arg Pro Thr Leu Gln Val Glu
115 120 125
Arg Arg His Pro Gly Leu Ser Ser Asp Ser Ala Ala Glu Ile Lys Lys
130 135 140
Arg Arg Lys Ala Ile Ala Ser Ser Asp Ser Glu Leu Glu Ser Leu Thr
145 150 155 160
Tyr Pro Asp Lys Pro Thr Lys Ala Asn Lys Arg Lys Val Ala Lys Glu
165 170 175
Ser Val Val Asp Ala Ser Glu Ser Asp Leu Asp Ser Ser Met Gln Ser
180 185 190
Ala Asp Glu Ser Thr Pro Gln Pro Leu Lys Ala Asn Gln Lys Pro Phe
195 200 205
Phe Pro Lys Val Phe Lys Lys Ile Lys Asp Ala Gly Lys Trp Val Arg
210 215 220
Asp Lys
225
<210> 73
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 73
Gln Asp Asn Lys Arg
1 5
<210> 74
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 74
Glu Cys Thr Gly Leu Ala Trp Glu Trp Trp Arg
1 5 10
<210> 75
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 75
Glu Ser Leu Leu Met Trp Ile Thr Gln Cys Arg
1 5 10
<210> 76
<211> 368
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 76
Met Val Thr Gly Trp His Arg Pro Thr Trp Ile Glu Ile Asp Arg Ala
1 5 10 15
Ala Ile Arg Glu Asn Ile Lys Asn Glu Gln Asn Lys Leu Pro Glu Ser
20 25 30
Val Asp Leu Trp Ala Val Val Lys Ala Asn Ala Tyr Gly His Gly Ile
35 40 45
Ile Glu Val Ala Arg Thr Ala Lys Glu Ala Gly Ala Lys Gly Phe Cys
50 55 60
Val Ala Ile Leu Asp Glu Ala Leu Ala Leu Arg Glu Ala Gly Phe Gln
65 70 75 80
Asp Asp Phe Ile Leu Val Leu Gly Ala Thr Arg Lys Glu Asp Ala Asn
85 90 95
Leu Ala Ala Lys Asn His Ile Ser Leu Thr Val Phe Arg Glu Asp Trp
100 105 110
Leu Glu Asn Leu Thr Leu Glu Ala Thr Leu Arg Ile His Leu Lys Val
115 120 125
Asp Ser Gly Met Gly Arg Leu Gly Ile Arg Thr Thr Glu Glu Ala Arg
130 135 140
Arg Ile Glu Ala Thr Ser Thr Asn Asp His Gln Leu Gln Leu Glu Gly
145 150 155 160
Ile Tyr Thr His Phe Ala Thr Ala Asp Gln Leu Glu Thr Ser Tyr Phe
165 170 175
Glu Gln Gln Leu Ala Lys Phe Gln Thr Ile Leu Thr Ser Leu Lys Lys
180 185 190
Arg Pro Thr Tyr Val His Thr Ala Asn Ser Ala Ala Ser Leu Leu Gln
195 200 205
Pro Gln Ile Gly Phe Asp Ala Ile Arg Phe Gly Ile Ser Met Tyr Gly
210 215 220
Leu Thr Pro Ser Thr Glu Ile Lys Thr Ser Leu Pro Phe Glu Leu Lys
225 230 235 240
Pro Ala Leu Ala Leu Tyr Thr Glu Met Val His Val Lys Glu Leu Ala
245 250 255
Pro Gly Asp Ser Val Ser Tyr Gly Ala Thr Tyr Thr Ala Thr Glu Arg
260 265 270
Glu Trp Val Ala Thr Leu Pro Ile Gly Tyr Ala Asp Gly Leu Ile Arg
275 280 285
His Tyr Ser Gly Phe His Val Leu Val Asp Gly Glu Pro Ala Pro Ile
290 295 300
Ile Gly Arg Val Cys Met Asp Gln Thr Ile Ile Lys Leu Pro Arg Glu
305 310 315 320
Phe Gln Thr Gly Ser Lys Val Thr Ile Ile Gly Lys Asp His Gly Asn
325 330 335
Thr Val Thr Ala Asp Asp Ala Ala Gln Tyr Leu Asp Thr Ile Asn Tyr
340 345 350
Glu Val Thr Cys Leu Leu Asn Glu Arg Ile Pro Arg Lys Tyr Ile His
355 360 365
<210> 77
<211> 289
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 77
Met Lys Val Leu Val Asn Asn His Leu Val Glu Arg Glu Asp Ala Thr
1 5 10 15
Val Asp Ile Glu Asp Arg Gly Tyr Gln Phe Gly Asp Gly Val Tyr Glu
20 25 30
Val Val Arg Leu Tyr Asn Gly Lys Phe Phe Thr Tyr Asn Glu His Ile
35 40 45
Asp Arg Leu Tyr Ala Ser Ala Ala Lys Ile Asp Leu Val Ile Pro Tyr
50 55 60
Ser Lys Glu Glu Leu Arg Glu Leu Leu Glu Lys Leu Val Ala Glu Asn
65 70 75 80
Asn Ile Asn Thr Gly Asn Val Tyr Leu Gln Val Thr Arg Gly Val Gln
85 90 95
Asn Pro Arg Asn His Val Ile Pro Asp Asp Phe Pro Leu Glu Gly Val
100 105 110
Leu Thr Ala Ala Ala Arg Glu Val Pro Arg Asn Glu Arg Gln Phe Val
115 120 125
Glu Gly Gly Thr Ala Ile Thr Glu Glu Asp Val Arg Trp Leu Arg Cys
130 135 140
Asp Ile Lys Ser Leu Asn Leu Leu Gly Asn Ile Leu Ala Lys Asn Lys
145 150 155 160
Ala His Gln Gln Asn Ala Leu Glu Ala Ile Leu His Arg Gly Glu Gln
165 170 175
Val Thr Glu Cys Ser Ala Ser Asn Val Ser Ile Ile Lys Asp Gly Val
180 185 190
Leu Trp Thr His Ala Ala Asp Asn Leu Ile Leu Asn Gly Ile Thr Arg
195 200 205
Gln Val Ile Ile Asp Val Ala Lys Lys Asn Gly Ile Pro Val Lys Glu
210 215 220
Ala Asp Phe Thr Leu Thr Asp Leu Arg Glu Ala Asp Glu Val Phe Ile
225 230 235 240
Ser Ser Thr Thr Ile Glu Ile Thr Pro Ile Thr His Ile Asp Gly Val
245 250 255
Gln Val Ala Asp Gly Lys Arg Gly Pro Ile Thr Ala Gln Leu His Gln
260 265 270
Tyr Phe Val Glu Glu Ile Thr Arg Ala Cys Gly Glu Leu Glu Phe Ala
275 280 285
Lys
<210> 78
<211> 1107
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 78
atggtgacag gctggcatcg tccaacatgg attgaaatag accgcgcagc aattcgcgaa 60
aatataaaaa atgaacaaaa taaactcccg gaaagtgtcg acttatgggc agtagtcaaa 120
gctaatgcat atggtcacgg aattatcgaa gttgctagga cggcgaaaga agctggagca 180
aaaggtttct gcgtagccat tttagatgag gcactggctc ttagagaagc tggatttcaa 240
gatgacttta ttcttgtgct tggtgcaacc agaaaagaag atgctaatct ggcagccaaa 300
aaccacattt cacttactgt ttttagagaa gattggctag agaatctaac gctagaagca 360
acacttcgaa ttcatttaaa agtagatagc ggtatggggc gtctcggtat tcgtacgact 420
gaagaagcac ggcgaattga agcaaccagt actaatgatc accaattaca actggaaggt 480
atttacacgc attttgcaac agccgaccag ctagaaacta gttattttga acaacaatta 540
gctaagttcc aaacgatttt aacgagttta aaaaaacgac caacttatgt tcatacagcc 600
aattcagctg cttcattgtt acagccacaa atcgggtttg atgcgattcg ctttggtatt 660
tcgatgtatg gattaactcc ctccacagaa atcaaaacta gcttgccgtt tgagcttaaa 720
cctgcacttg cactctatac cgagatggtt catgtgaaag aacttgcacc aggcgatagc 780
gttagctacg gagcaactta tacagcaaca gagcgagaat gggttgcgac attaccaatt 840
ggctatgcgg atggattgat tcgtcattac agtggtttcc atgttttagt agacggtgaa 900
ccagctccaa tcattggtcg agtttgtatg gatcaaacca tcataaaact accacgtgaa 960
tttcaaactg gttcaaaagt aacgataatt ggcaaagatc atggtaacac ggtaacagca 1020
gatgatgccg ctcaatattt agatacaatt aattatgagg taacttgttt gttaaatgag 1080
cgcataccta gaaaatacat ccattag 1107
<210> 79
<211> 870
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 79
atgaaagtat tagtaaataa ccatttagtt gaaagagaag atgccacagt tgacattgaa 60
gaccgcggat atcagtttgg tgatggtgta tatgaagtag ttcgtctata taatggaaaa 120
ttctttactt ataatgaaca cattgatcgc ttatatgcta gtgcagcaaa aattgactta 180
gttattcctt attccaaaga agagctacgt gaattacttg aaaaattagt tgccgaaaat 240
aatatcaata cagggaatgt ctatttacaa gtgactcgtg gtgttcaaaa cccacgtaat 300
catgtaatcc ctgatgattt ccctctagaa ggcgttttaa cagcagcagc tcgtgaagta 360
cctagaaacg agcgtcaatt cgttgaaggt ggaacggcga ttacagaaga agatgtgcgc 420
tggttacgct gtgatattaa gagcttaaac cttttaggaa atattctagc aaaaaataaa 480
gcacatcaac aaaatgcttt ggaagctatt ttacatcgcg gggaacaagt aacagaatgt 540
tctgcttcaa acgtttctat tattaaagat ggtgtattat ggacgcatgc ggcagataac 600
ttaatcttaa atggtatcac tcgtcaagtt atcattgatg ttgcgaaaaa gaatggcatt 660
cctgttaaag aagcggattt cactttaaca gaccttcgtg aagcggatga agtgttcatt 720
tcaagtacaa ctattgaaat tacacctatt acgcatattg acggagttca agtagctgac 780
ggaaaacgtg gaccaattac agcgcaactt catcaatatt ttgtagaaga aatcactcgt 840
gcatgtggcg aattagagtt tgcaaaataa 870
<210> 80
<211> 237
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 80
Met Asn Ala Gln Ala Glu Glu Phe Lys Lys Tyr Leu Glu Thr Asn Gly
1 5 10 15
Ile Lys Pro Lys Gln Phe His Lys Lys Glu Leu Ile Phe Asn Gln Trp
20 25 30
Asp Pro Gln Glu Tyr Cys Ile Phe Leu Tyr Asp Gly Ile Thr Lys Leu
35 40 45
Thr Ser Ile Ser Glu Asn Gly Thr Ile Met Asn Leu Gln Tyr Tyr Lys
50 55 60
Gly Ala Phe Val Ile Met Ser Gly Phe Ile Asp Thr Glu Thr Ser Val
65 70 75 80
Gly Tyr Tyr Asn Leu Glu Val Ile Ser Glu Gln Ala Thr Ala Tyr Val
85 90 95
Ile Lys Ile Asn Glu Leu Lys Glu Leu Leu Ser Lys Asn Leu Thr His
100 105 110
Phe Phe Tyr Val Phe Gln Thr Leu Gln Lys Gln Val Ser Tyr Ser Leu
115 120 125
Ala Lys Phe Asn Asp Phe Ser Ile Asn Gly Lys Leu Gly Ser Ile Cys
130 135 140
Gly Gln Leu Leu Ile Leu Thr Tyr Val Tyr Gly Lys Glu Thr Pro Asp
145 150 155 160
Gly Ile Lys Ile Thr Leu Asp Asn Leu Thr Met Gln Glu Leu Gly Tyr
165 170 175
Ser Ser Gly Ile Ala His Ser Ser Ala Val Ser Arg Ile Ile Ser Lys
180 185 190
Leu Lys Gln Glu Lys Val Ile Val Tyr Lys Asn Ser Cys Phe Tyr Val
195 200 205
Gln Asn Leu Asp Tyr Leu Lys Arg Tyr Ala Pro Lys Leu Asp Glu Trp
210 215 220
Phe Tyr Leu Ala Cys Pro Ala Thr Trp Gly Lys Leu Asn
225 230 235
<210> 81
<211> 714
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 81
atgaacgctc aagcagaaga attcaaaaaa tatttagaaa ctaacgggat aaaaccaaaa 60
caatttcata aaaaagaact tatttttaac caatgggatc cacaagaata ttgtattttt 120
ctatatgatg gtatcacaaa gctcacgagt attagcgaga acgggaccat catgaattta 180
caatactaca aaggggcttt cgttataatg tctggcttta ttgatacaga aacatcggtt 240
ggctattata atttagaagt cattagcgag caggctaccg catacgttat caaaataaac 300
gaactaaaag aactactgag caaaaatctt acgcactttt tctatgtttt ccaaacccta 360
caaaaacaag tttcatacag cctagctaaa tttaatgatt tttcgattaa cgggaagctt 420
ggctctattt gcggtcaact tttaatcctg acctatgtgt atggtaaaga aactcctgat 480
ggcatcaaga ttacactgga taatttaaca atgcaggagt taggatattc aagtggcatc 540
gcacatagct cagctgttag cagaattatt tccaaattaa agcaagagaa agttatcgtg 600
tataaaaatt catgctttta tgtacaaaat cttgattatc tcaaaagata tgcccctaaa 660
ttagatgaat ggttttattt agcatgtcct gctacttggg gaaaattaaa ttaa 714
<210> 82
<211> 237
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 82
Met Asn Ala Gln Ala Glu Glu Phe Lys Lys Tyr Leu Glu Thr Asn Gly
1 5 10 15
Ile Lys Pro Lys Gln Phe His Lys Lys Glu Leu Ile Phe Asn Gln Trp
20 25 30
Asp Pro Gln Glu Tyr Cys Ile Phe Leu Tyr Asp Gly Ile Thr Lys Leu
35 40 45
Thr Ser Ile Ser Glu Asn Gly Thr Ile Met Asn Leu Gln Tyr Tyr Lys
50 55 60
Gly Ala Phe Val Ile Met Ser Gly Phe Ile Asp Thr Glu Thr Ser Val
65 70 75 80
Gly Tyr Tyr Asn Leu Glu Val Ile Ser Glu Gln Ala Thr Ala Tyr Val
85 90 95
Ile Lys Ile Asn Glu Leu Lys Glu Leu Leu Ser Lys Asn Leu Thr His
100 105 110
Phe Phe Tyr Val Phe Gln Thr Leu Gln Lys Gln Val Ser Tyr Ser Leu
115 120 125
Ala Lys Phe Asn Val Phe Ser Ile Asn Gly Lys Leu Gly Ser Ile Cys
130 135 140
Gly Gln Leu Leu Ile Leu Thr Tyr Val Tyr Gly Lys Glu Thr Pro Asp
145 150 155 160
Gly Ile Lys Ile Thr Leu Asp Asn Leu Thr Met Gln Glu Leu Gly Tyr
165 170 175
Ser Ser Gly Ile Ala His Ser Ser Ala Val Ser Arg Ile Ile Ser Lys
180 185 190
Leu Lys Gln Glu Lys Val Ile Val Tyr Lys Asn Ser Cys Phe Tyr Val
195 200 205
Gln Asn Arg Asp Tyr Leu Lys Arg Tyr Ala Pro Lys Leu Asp Glu Trp
210 215 220
Phe Tyr Leu Ala Cys Pro Ala Thr Trp Gly Lys Leu Asn
225 230 235
<210> 83
<211> 713
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 83
atgaacgctc aagcagaaga attcaaaaaa tatttagaaa ctaacgggat aaaaccaaaa 60
caatttcata aaaaagaact tatttttaac caatgggatc cacaagaata ttgtattttt 120
ctatatgatg gtatcacaaa gctcacgagt attagcgaga acgggaccat catgaattta 180
caatactaca aaggggcttt cgttataatg tctggcttta ttgatacaga aacatcggtt 240
ggctattata atttagaagt cattagcgag caggctaccg catacgttat caaaataaac 300
gaactaaaag aactactgag caaaaatctt acgcactttt tctatgtttt ccaaacccta 360
caaaaacaag tttcatacag cctagctaaa tttaatgttt tttcgattaa cgggaagctt 420
ggctctattt gcggtcaact tttaatcctg acctatgtgt atggtaaaga aactcctgat 480
ggcatcaaga ttacactgga taatttaaca atgcaggagt taggatattc aagtggcatc 540
gcacatagct cagctgttag cagaattatt tccaaattaa agcaagagaa agttatcgtg 600
tataaaaatt catgctttta tgtacaaaat ctgattatct caaaagatat gcccctaaat 660
tagatgaatg gttttattta gcatgtcctg ctacttgggg aaaattaaat taa 713
<210> 84
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 84
ggtggtggag ga 12
<210> 85
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 85
ggtggaggtg ga 12
<210> 86
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 86
ggtggaggag gt 12
<210> 87
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 87
ggaggtggtg ga 12
<210> 88
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 88
ggaggaggtg gt 12
<210> 89
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 89
ggaggtggag gt 12
<210> 90
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 90
ggaggaggag gt 12
<210> 91
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 91
ggaggaggtg ga 12
<210> 92
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 92
ggaggtggag ga 12
<210> 93
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 93
ggtggaggag ga 12
<210> 94
<211> 12
<212> DNA
<213> 人工序列
<220>
<223> 合成
<400> 94
ggaggaggag ga 12
<210> 95
<211> 529
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 95
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Lys Asp Ala Ser Ala Phe Asn Lys
20 25 30
Glu Asn Ser Ile Ser Ser Met Ala Pro Pro Ala Ser Pro Pro Ala Ser
35 40 45
Pro Lys Thr Pro Ile Glu Lys Lys His Ala Asp Glu Ile Asp Lys Tyr
50 55 60
Ile Gln Gly Leu Asp Tyr Asn Lys Asn Asn Val Leu Val Tyr His Gly
65 70 75 80
Asp Ala Val Thr Asn Val Pro Pro Arg Lys Gly Tyr Lys Asp Gly Asn
85 90 95
Glu Tyr Ile Val Val Glu Lys Lys Lys Lys Ser Ile Asn Gln Asn Asn
100 105 110
Ala Asp Ile Gln Val Val Asn Ala Ile Ser Ser Leu Thr Tyr Pro Gly
115 120 125
Ala Leu Val Lys Ala Asn Ser Glu Leu Val Glu Asn Gln Pro Asp Val
130 135 140
Leu Pro Val Lys Arg Asp Ser Leu Thr Leu Ser Ile Asp Leu Pro Gly
145 150 155 160
Met Thr Asn Gln Asp Asn Lys Ile Val Val Lys Asn Ala Thr Lys Ser
165 170 175
Asn Val Asn Asn Ala Val Asn Thr Leu Val Glu Arg Trp Asn Glu Lys
180 185 190
Tyr Ala Gln Ala Tyr Pro Asn Val Ser Ala Lys Ile Asp Tyr Asp Asp
195 200 205
Glu Met Ala Tyr Ser Glu Ser Gln Leu Ile Ala Lys Phe Gly Thr Ala
210 215 220
Phe Lys Ala Val Asn Asn Ser Leu Asn Val Asn Phe Gly Ala Ile Ser
225 230 235 240
Glu Gly Lys Met Gln Glu Glu Val Ile Ser Phe Lys Gln Ile Tyr Tyr
245 250 255
Asn Val Asn Val Asn Glu Pro Thr Arg Pro Ser Arg Phe Phe Gly Lys
260 265 270
Ala Val Thr Lys Glu Gln Leu Gln Ala Leu Gly Val Asn Ala Glu Asn
275 280 285
Pro Pro Ala Tyr Ile Ser Ser Val Ala Tyr Gly Arg Gln Val Tyr Leu
290 295 300
Lys Leu Ser Thr Asn Ser His Ser Thr Lys Val Lys Ala Ala Phe Asp
305 310 315 320
Ala Ala Val Ser Gly Lys Ser Val Ser Gly Asp Val Glu Leu Thr Asn
325 330 335
Ile Ile Lys Asn Ser Ser Phe Lys Ala Val Ile Tyr Gly Gly Ser Ala
340 345 350
Lys Asp Glu Val Gln Ile Ile Asp Gly Asn Leu Gly Asp Leu Arg Asp
355 360 365
Ile Leu Lys Lys Gly Ala Thr Phe Asn Arg Glu Thr Pro Gly Val Pro
370 375 380
Ile Ala Tyr Thr Thr Asn Phe Leu Lys Asp Asn Glu Leu Ala Val Ile
385 390 395 400
Lys Asn Asn Ser Glu Tyr Ile Glu Thr Thr Ser Lys Ala Tyr Thr Asp
405 410 415
Gly Lys Ile Asn Ile Asp His Ser Gly Gly Tyr Val Ala Gln Phe Asn
420 425 430
Ile Ser Trp Asp Glu Val Asn Tyr Asp Pro Glu Gly Asn Glu Ile Val
435 440 445
Gln His Lys Asn Trp Ser Glu Asn Asn Lys Ser Lys Leu Ala His Phe
450 455 460
Thr Ser Ser Ile Tyr Leu Pro Gly Asn Ala Arg Asn Ile Asn Val Tyr
465 470 475 480
Ala Lys Glu Ala Thr Gly Leu Ala Trp Glu Ala Ala Arg Thr Val Ile
485 490 495
Asp Asp Arg Asn Leu Pro Leu Val Lys Asn Arg Asn Ile Ser Ile Trp
500 505 510
Gly Thr Thr Leu Tyr Pro Lys Tyr Ser Asn Lys Val Asp Asn Pro Ile
515 520 525
Glu
<210> 96
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 96
Glu Ala Thr Gly Leu Ala Trp Glu Ala Ala Arg
1 5 10
<210> 97
<211> 25
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 97
Met Lys Lys Ile Met Leu Val Phe Ile Thr Leu Ile Leu Val Ser Leu
1 5 10 15
Pro Ile Ala Gln Gln Thr Glu Ala Lys
20 25
<210> 98
<211> 29
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 98
Met Gly Leu Asn Arg Phe Met Arg Ala Met Met Val Val Phe Ile Thr
1 5 10 15
Ala Asn Cys Ile Thr Ile Asn Pro Asp Ile Ile Phe Ala
20 25
<210> 99
<211> 21
<212> PRT
<213> 人工序列
<220>
<223> 合成
<400> 99
Asp Tyr Lys Asp His Asp Gly Asp Tyr Lys Asp His Asp Ile Asp Tyr
1 5 10 15
Lys Asp Asp Asp Lys
20
Claims (18)
1.一种评估测试李斯特菌菌株的减毒或感染性的方法,所述方法包括:
(a)用所述测试李斯特菌菌株感染分化的THP-1细胞,其中在用所述测试李斯特菌菌株感染之前,已将所述THP-1细胞分化为巨噬细胞;
(b)裂解所述THP-1细胞,并将裂解物平板接种在琼脂上;以及
(c)通过在所述琼脂上的生长来计数已在所述THP-1细胞内部繁殖的所述李斯特菌。
2.根据权利要求1所述的方法,其进一步包括在步骤(a)之前,使用佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA)将所述THP-1细胞分化为巨噬细胞。
3.根据前述权利要求中任一项所述的方法,其中步骤(a)包括以1:1的感染复数(MOI)感染所述分化的THP-1细胞。
4.根据前述权利要求中任一项所述的方法,其进一步包括在步骤(a)与(b)之间,杀死未被所述THP-1细胞摄取的李斯特菌。
5.根据权利要求4所述的方法,其中使用抗生素进行所述杀死,任选地其中所述抗生素是庆大霉素。
6.根据前述权利要求中任一项所述的方法,其中在感染后0小时进行步骤(b)。
7.根据前述权利要求中任一项所述的方法,其中在感染后3小时进行步骤(b)。
8.根据前述权利要求中任一项所述的方法,其进一步包括比较所述测试李斯特菌菌株与野生型李斯特菌菌株和/或参考样品的摄取和细胞内生长。
9.根据前述权利要求中任一项所述的方法,其中所述测试李斯特菌菌株是单核细胞增生性李斯特菌菌株。
10.根据前述权利要求中任一项所述的方法,其中所述测试李斯特菌菌株是重组李斯特菌菌株,所述重组李斯特菌菌株包括有包括对融合多肽进行编码的第一开放阅读框的核酸,其中所述融合多肽包括与疾病相关型抗原肽融合的含PEST的肽。
11.根据权利要求10所述的方法,其中所述含PEST的肽是李斯特菌溶血素O(LLO)或其片段,并且所述疾病相关型抗原肽是人乳头瘤病毒(HPV)蛋白E7或其片段。
12.根据权利要求10或11所述的方法,其中所述重组李斯特菌菌株是减毒的单核细胞增生性李斯特菌菌株,所述减毒的单核细胞增生性李斯特菌菌株包括prfA的缺失或prfA中的失活突变,其中所述核酸位于附加型质粒中并且包括对D133V PrfA突变蛋白进行编码的第二开放阅读框。
13.根据权利要求10所述的方法,其中所述重组李斯特菌菌株是减毒的单核细胞增生性李斯特菌菌株,所述减毒的单核细胞增生性李斯特菌菌株包括actA、dal和dat的缺失或actA、dal和dat中的失活突变,其中所述核酸位于附加型质粒中并且包括对丙氨酸消旋酶或D-氨基酸氨基转移酶进行编码的第二开放阅读框,并且其中所述含PEST的肽是李斯特菌溶血素O(LLO)的N端片段。
14.一种评估测试细菌菌株的减毒或感染性的方法,所述方法包括:
(a)分化THP-1细胞;
(b)用所述测试细菌菌株感染分化的THP-1细胞,其中所述感染包括:
(i)用所述测试细菌菌株接种所述分化的THP-1细胞;
(ii)将所述测试细菌菌株与所述分化的THP-1细胞一起孵育1-5小时,以形成感染的THP1细胞;
(iii)从所述感染的THP-1细胞中去除细胞外细菌;以及
(iv)将所述感染的THP-1细胞在生长培养基中孵育0-10小时;
(c)裂解所述感染的THP-1细胞以形成裂解物;
(d)将所述裂解物或所述裂解物的稀释液平板接种在含有能够支持所述细菌生长的培养基的板上;以及
(e)计数所述板上的所述细菌的菌落形成单位。
15.根据权利要求14所述的方法,其中感染所述分化的THP-1细胞的步骤是以1:1的感染复数(MOI)进行的。
16.根据权利要求14或15所述的方法,其中去除细胞外细菌的步骤包括添加对所述细菌有效的抗生素,任选地其中所述抗生素是庆大霉素。
17.根据权利要求14到16中任一项所述的方法,其中将所述感染的THP-1细胞在生长培养基中孵育0小时、1小时、3小时或5小时。
18.根据权利要求14到17中任一项所述的方法,其中所述测试细菌菌株是单核细胞增生性李斯特菌菌株。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201862640855P | 2018-03-09 | 2018-03-09 | |
| US62/640,855 | 2018-03-09 | ||
| PCT/US2019/021303 WO2019173684A1 (en) | 2018-03-09 | 2019-03-08 | Compositions and methods for evaluating attenuation and infectivity of listeria strains |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN112074737A true CN112074737A (zh) | 2020-12-11 |
Family
ID=66102746
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201980029910.3A Pending CN112074737A (zh) | 2018-03-09 | 2019-03-08 | 用于评估李斯特菌菌株的减毒和感染性的组合物和方法 |
Country Status (11)
| Country | Link |
|---|---|
| US (1) | US20210003558A1 (zh) |
| EP (1) | EP3762719A1 (zh) |
| JP (2) | JP2021516972A (zh) |
| KR (1) | KR20200130399A (zh) |
| CN (1) | CN112074737A (zh) |
| AU (1) | AU2019231783B2 (zh) |
| CA (1) | CA3093467C (zh) |
| IL (1) | IL277255A (zh) |
| MX (1) | MX2020009405A (zh) |
| SG (1) | SG11202008820WA (zh) |
| WO (1) | WO2019173684A1 (zh) |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US9012141B2 (en) | 2000-03-27 | 2015-04-21 | Advaxis, Inc. | Compositions and methods comprising KLK3 of FOLH1 antigen |
| MA41644A (fr) | 2015-03-03 | 2018-01-09 | Advaxis Inc | Compositions à base de listeria comprenant un système d'expression de minigènes codant pour des peptides, et leurs procédés d'utilisation |
| SG10202105561PA (en) | 2016-11-30 | 2021-07-29 | Advaxis Inc | Immunogenic compositions targeting recurrent cancer mutations and methods of use thereof |
| KR20200044982A (ko) | 2017-09-19 | 2020-04-29 | 어드박시스, 인크. | 박테리아 또는 리스테리아 균주의 동결건조를 위한 조성물 및 방법 |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2017048714A1 (en) * | 2015-09-15 | 2017-03-23 | Advaxis, Inc. | Manufacturing method of an immunotherapeutic formulation comprising a recombinant listeria strain |
| CN107073094A (zh) * | 2014-07-18 | 2017-08-18 | 阿德瓦希斯公司 | 表达异源性抗原融合蛋白的重组李斯特菌菌株及其使用方法 |
Family Cites Families (19)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4751180A (en) | 1985-03-28 | 1988-06-14 | Chiron Corporation | Expression using fused genes providing for protein product |
| US4935233A (en) | 1985-12-02 | 1990-06-19 | G. D. Searle And Company | Covalently linked polypeptide cell modulators |
| US7635479B2 (en) | 2000-03-29 | 2009-12-22 | The Trustees Of The University Of Pennsylvania | Composition and methods for enhancing immunogenecity of antigens |
| US8114414B2 (en) | 1994-11-08 | 2012-02-14 | The Trustees Of The University Of Pennsylvania | Compositions and methods for treatment of cervical cancer |
| US6099848A (en) | 1997-11-18 | 2000-08-08 | The Trustees Of The University Of Pennsylvania | Immunogenic compositions comprising DAL/DAT double-mutant, auxotrophic, attenuated strains of Listeria and their methods of use |
| US6855320B2 (en) | 2000-03-29 | 2005-02-15 | The Trustees Of The University Of Pennsylvania | Fusion of non-hemolytic, truncated form of listeriolysin O to antigens to enhance immunogenicity |
| US8771702B2 (en) | 2001-03-26 | 2014-07-08 | The Trustees Of The University Of Pennsylvania | Non-hemolytic LLO fusion proteins and methods of utilizing same |
| EP1513924A4 (en) | 2002-05-29 | 2008-05-21 | Univ California | ATTENUATED LISTERIA SPP. AND METHOD OF USE THEREOF |
| JP4545151B2 (ja) | 2003-02-06 | 2010-09-15 | シーラス コーポレイション | 非食細胞中への侵入について減弱化されているリステリア、そのリステリアを含むワクチン、およびそれらの使用法 |
| US7935804B2 (en) | 2006-03-01 | 2011-05-03 | Aduro Biotech | Engineered Listeria and methods of use thereof |
| US7665238B2 (en) | 2006-04-03 | 2010-02-23 | S.C. Johnson & Son, Inc. | Air freshener with holder |
| EP2085466A1 (en) * | 2008-01-29 | 2009-08-05 | AEterna Zentaris GmbH | Non-pathogenic and/or attenuated bacteria capable of inducing apoptosis in macrophages, process of manufacturing and uses thereof |
| WO2011060260A2 (en) | 2009-11-11 | 2011-05-19 | Advaxis | Compositions and methods for prevention of escape mutation in the treatment of her2/neu over-expressing tumors |
| ES2636902T3 (es) | 2012-05-25 | 2017-10-10 | The Regents Of The University Of California | Métodos y composiciones para la modificación de ADN objetivo dirigida por ARN y para la modulación de la transcripción dirigida por ARN |
| SG10201700916SA (en) | 2012-12-27 | 2017-03-30 | Aduro Biotech Inc | Signal peptide fusion partners facilitating listerial expression of antigenic sequences and methods of preparation and use thereof |
| SG10202011841WA (en) * | 2014-04-24 | 2021-01-28 | Advaxis Inc | Recombinant listeria vaccine strains and methods of producing the same |
| MA41218A (fr) * | 2014-12-19 | 2017-10-24 | Advaxis Inc | Combinaison de vaccin à base de listeria comportant des anticorps anti-ox40 ou anti-gitr |
| MA42263A (fr) * | 2015-06-24 | 2021-03-31 | Advaxis Inc | Dispositif de fabrication et procédé pour immunothérapie fondée sur un vecteur d'administration personnalisé |
| WO2018129306A1 (en) * | 2017-01-05 | 2018-07-12 | Advaxis, Inc. | Recombinant listeria vaccine strains and methods of using the same in cancer immunotherapy |
-
2019
- 2019-03-08 CN CN201980029910.3A patent/CN112074737A/zh active Pending
- 2019-03-08 CA CA3093467A patent/CA3093467C/en active Active
- 2019-03-08 JP JP2020547226A patent/JP2021516972A/ja active Pending
- 2019-03-08 AU AU2019231783A patent/AU2019231783B2/en not_active Expired - Fee Related
- 2019-03-08 KR KR1020207028931A patent/KR20200130399A/ko active Search and Examination
- 2019-03-08 MX MX2020009405A patent/MX2020009405A/es unknown
- 2019-03-08 WO PCT/US2019/021303 patent/WO2019173684A1/en active Application Filing
- 2019-03-08 SG SG11202008820WA patent/SG11202008820WA/en unknown
- 2019-03-08 US US16/979,436 patent/US20210003558A1/en not_active Abandoned
- 2019-03-08 EP EP19717010.3A patent/EP3762719A1/en not_active Withdrawn
-
2020
- 2020-09-09 IL IL277255A patent/IL277255A/en unknown
-
2023
- 2023-10-06 JP JP2023174128A patent/JP2023168614A/ja active Pending
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN107073094A (zh) * | 2014-07-18 | 2017-08-18 | 阿德瓦希斯公司 | 表达异源性抗原融合蛋白的重组李斯特菌菌株及其使用方法 |
| WO2017048714A1 (en) * | 2015-09-15 | 2017-03-23 | Advaxis, Inc. | Manufacturing method of an immunotherapeutic formulation comprising a recombinant listeria strain |
Non-Patent Citations (3)
| Title |
|---|
| AMRITA LAMA等: "Identification of Conserved ABC Importers Necessary for Intracellular Survival of Legionella pneumophila in Multiple Hosts", FRONTIERS IN CELLULAR AND INFECTION MICROBIOLOGY, 30 November 2017 (2017-11-30), pages 1 - 2 * |
| MARIA PIA CONTE等: "Effect of Acid Adaptation on the Fate of Listeria monocytogenes inTHP-1 Human Macrophages Activated by Gamma Interferon", INFECTION AND IMMUNITY, 31 August 2002 (2002-08-31), pages 3 - 4 * |
| NRUSINGH P. MOHAPATRA等: "Identification of an Orphan Response Regulator Required for the Virulence of Francisella spp. and Transcription of Pathogenicity Island Genes", INFECTION AND IMMUNITY, 30 June 2007 (2007-06-30), pages 3307 * |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2019173684A1 (en) | 2019-09-12 |
| JP2023168614A (ja) | 2023-11-24 |
| IL277255A (en) | 2020-10-29 |
| CA3093467A1 (en) | 2019-09-12 |
| AU2019231783B2 (en) | 2023-11-16 |
| EP3762719A1 (en) | 2021-01-13 |
| JP2021516972A (ja) | 2021-07-15 |
| AU2019231783A1 (en) | 2020-10-08 |
| SG11202008820WA (en) | 2020-10-29 |
| MX2020009405A (es) | 2021-01-08 |
| US20210003558A1 (en) | 2021-01-07 |
| CA3093467C (en) | 2022-12-06 |
| KR20200130399A (ko) | 2020-11-18 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| AU2019231783B2 (en) | Compositions and methods for evaluating attenuation and infectivity of Listeria strains | |
| Divangahi et al. | NOD2-deficient mice have impaired resistance to Mycobacterium tuberculosis infection through defective innate and adaptive immunity | |
| Brockstedt et al. | Killed but metabolically active microbes: a new vaccine paradigm for eliciting effector T-cell responses and protective immunity | |
| KR20190082850A (ko) | 반복되는 암 돌연변이를 표적화하는 면역원 조성물 및 그것의 사용 방법 | |
| JP5713523B2 (ja) | エンドソームエスケープ能力増強組み換えbcg株類 | |
| KR20180026670A (ko) | 맞춤형 전달 벡터-기반의 면역 요법 및 이의 용도 | |
| US20210239681A1 (en) | Compositions and methods for evaluating potency of listeria-based immunotherapeutics | |
| CN111655714A (zh) | 来自癌症相关蛋白的免疫原性异变肽及其使用方法 | |
| CN107073094A (zh) | 表达异源性抗原融合蛋白的重组李斯特菌菌株及其使用方法 | |
| CN111356760A (zh) | 细菌或李斯特菌菌株的冻干组合物和方法 | |
| KR20140134695A (ko) | 리스테리아 백신 치료 후 억제 세포 기능 저해 | |
| CN109641945A (zh) | 包含维尔姆斯瘤蛋白抗原的基于李斯特菌的免疫原性组合物及其使用方法 | |
| TW201833323A (zh) | 重組李斯特菌屬疫苗菌株及在癌症免疫治療中使用該菌株之方法 | |
| Verch et al. | Listeria monocytogenes-based antibiotic resistance gene-free antigen delivery system applicable to other bacterial vectors and DNA vaccines | |
| US11969468B2 (en) | Live self-destructing bacterial adjuvants to enhance induction of immunity | |
| Roider et al. | Invasion and destruction of a murine fibrosarcoma by Salmonella-induced effector CD8 T cells as a therapeutic intervention against cancer | |
| Michell et al. | Deletion of the Bacillus anthracis capB homologue in Francisella tularensis subspecies tularensis generates an attenuated strain that protects mice against virulent tularaemia | |
| US20170173137A1 (en) | Prevention and Treatment of Mycobacterium Infection | |
| Yu et al. | A method to generate recombinant Salmonella typhi Ty21a strains expressing multiple heterologous genes using an improved recombineering strategy | |
| WO2003079792A1 (en) | Methods and compositions for vaccination against or involving enterobacteriaceae bacteria | |
| Tzelepis et al. | Modulation of antigenic location converts chronic into acute infection by forcing CD8+ T cell recognition | |
| Sizemore et al. | Interaction of Salmonella typhi strains with cultured human monocyte-derived macrophages | |
| US20230165955A1 (en) | Live self-destructing bacterial adjuvants to enhance induction of immunity | |
| Shaku | Peptidoglycan amidation enzymes in Mycobacterium tuberculosis are new drug targets for Tuberculosis treatment and development of a novel TB vaccine | |
| Iwanowitsch et al. | The JMU-SalVac-System: A Novel, Versatile Approach to Oral Live Vaccine Development |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |