
Lobopodia
Pour les pseudopodes, voir lobopode.
Lobopodiens

| Règne | Animalia |
|---|---|
| Sous-règne | Eumetazoa |
| Clade | Bilateria |
| Clade | Nephrozoa |
| Super-embr. | Ecdysozoa |
| Clade | Panarthropoda |
- Lobopoda Cavalier-Smith (1998)
Les lobopodiens, membres du groupe informel Lobopodia[1] (du grec, signifiant « pied pataud »), ou de l'ancien embranchement Lobopoda Cavalier-Smith (1998)[2], sont des panarthropodes possédant des pattes trapues appelées lobopodes[3], terme aussi utilisé comme nom pour le groupe tout entier[4],[5]. Bien que la définition des lobopodiens varie selon les auteurs, le terme renvoie habituellement à un groupe de panarthropodes vermiformes à corps mous fossiles comme Aysheaia et Hallucigenia[3].
Le plus ancien fossile de lobopodiens à peu-près complet date du Cambrien inférieur ; on en connaît dans des Lagerstätten de l'Ordovicien, du Silurien et du Carbonifère[6],[7],[8]. Certains présentent des griffes, des plaques ou des épines dures, qui sont couramment conservées comme microfossiles carbonés ou minéralisés dans les strates cambriennes[9],[10]
Définition
[modifier | modifier le code]
L'extension du concept de Lobopodien diffère selon les auteurs. Dans son sens le plus fréquent et le plus limité, il renvoie à un taxon de panarthropodes vermiformes possédant des lobopodes, principalement du Cambrien — par exemple Aysheaia, Hallucigenia et Xenusion (en), traditionnellement réunis dans la classe des xenusides (en). Les genres de dinocaridides Pambdelurion et Kerygmachela peuvent aussi être considérés comme des lobopodiens[11],[12], sous le nom de « lobopodiens à branchies[13] ». Selon cette définition, « Lobopodia » n'est composé que de taxons éteints et est largement accepté comme un groupe informel, paraphylétique, coiffant trois embranchements des panarthropodes : Onychophora (les vers de velours), Tardigrada (les oursons d'eau) et Arthropoda (les arthropodes)[3],[14],[15],[16],[17],[18].
Une définition plus large des lobopodiens y intègrerait aussi Onychophora et Tardigrada[19],[11],[2], qui possèdent aussi des pattes lobopodiennes[3]. « Lobopodia » peut aussi renvoyer à un clade-frère des arthropodes qui ne serait composé que de Tardigrada et Onychophora[20]. Lobopodia comprend aussi parfois Pentastomida, un groupe de panarthropodes parasites qui s'est révélé être un taxon de Crustacés hautement spécialisés[21],[22]. La définition la plus large propose un super-embranchement monophylétique Lobopodia qui serait équivalent à Panarthropoda[23],[4].
Taxons représentatifs
[modifier | modifier le code]

Les genres les plus connus sont Aysheaia, découvert au Canada dans les schistes de Burgess, et Hallucigenia, connu à la fois dans les schistes de Maotianshan au Yunnan et dans les schistes de Burgess. Aysheaia pedunculata possède apparemment une morphologie basale pour un lobopodien[14] — une cuticule nettement annelée, une ouverture buccale à une extrémité, des appendices frontaux spécialisés et des lobopodes trapus terminés par des griffes. Hallucigenia sparsa est devenu célèbre pour l'histoire de son interprétation — il a d'abord été reconstruit avec de longues pattes comme des échasses et de mystérieuses protubérances dorsales, et longtemps considéré comme un exemple des morphologies bizarres expérimentées par la nature au cours du Cambrien[24]. D'autres découvertes ont ensuite montré que cette reconstruction retournait l'animal : en interprétant les « échasses » comme des pointes dorsales, les protubérances « dorsales » charnues deviennent des lobopodes allongés. Des reconstructions plus récentes ont même inversé l'avant et l'arrière de l'animal : la trace bulbeuse interprétée comme une tête était en fait le contenu de son tube digestif expulsé par l'anus[9],[15].
Microdictyon est un autre genre célèbre ressemblant à Hallucigenia, mais au lieu d'épines, il a des paires de plaques, souvent retrouvées désarticulées parmi les petits fossiles coquilliers (SSF). Xenusion (en) est le fossile le plus ancien connu parmi les lobopodiens décrits, remontant à l'étage 2 du Cambrien[19],[12]. Luolishania est un exemple classique de lobopodien possédant de multiples paires d'appendices spécialisés[25]. Les lobopodiens à branchies Kerygmachela et Pambdelurion mettent en lumière les relations entre ce groupe et les arthropodes, car ils possèdent à la fois des affinités avec les lobopodiens et des caractères liés au groupe-racine des arthropodes[13],[23].
Morphologie
[modifier | modifier le code]-
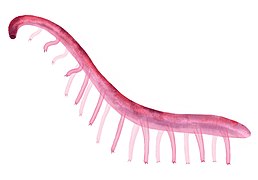 Restitution paléoartistique de Paucipodia inermis.
Restitution paléoartistique de Paucipodia inermis. -



La plupart des lobopodiens ne dépassent pas quelques centimètres de long, mais certains genres ont dépassé 20 cm[5]. Leur corps est annelé, même si les anneaux sont parfois difficiles à voir en raison de leur finesse (~0,2 mm) et du manque de relief des fossiles[26]. le corps et les appendices sont de section circulaire[26].
Lobopodes
[modifier | modifier le code]
Les membres des lobopodiens, appelés lobopodes[3] ou membres lobopodiens[11], sont grossièrement coniques, s'effilant du corps jusqu'à leur extrémité griffue[26],[14] ou non-griffue[5],[27],[28],[12]. Les pattes les plus longues et les plus robustes sont habituellement celles du milieu du tronc, celles de l'avant et de l'arrière étant plus grêles[26]. Les griffes, quand il y en a, sont légèrement courbes et leur longueur est grossièrement proportionnelle à celle du membre qui les porte[26]. Dans certains genres, les lobopodes peuvent porter des structures supplémentaires comme des épines (par exemple Diania[28]), des excroissances charnues (par exemple Onychodictyon), ou des tubercules (par exemple Jianshanopodia[5]). Il n'y a aucun signe d'arthropodisation (développement d'un exosquelette dur et segmentation des appendices des panarthropodes) chez les lobopodiens connus, même pour ceux qui appartiennent au groupe-racine des arthropodes (par exemple les lobopodiens à branchies et les siberides), et l'arthropodisation soupçonnée des membres de Diania[29] est considérée comme une mauvaise interprétation[28],[12].
Tête
[modifier | modifier le code]
La tête est plus ou moins bulbeuse[3] et peut porter une paire d'appendices pré-oculaires protocérébraux[17] – par exemple des antennes primaires[27],[25],[17],[12] ou des appendices frontaux bien développés[3],[11],[30],[5] bien distincts des lobopodes du tronc[17],[31] (à l'exception d’Antennacanthopodia, qui a deux paires d'appendices sur la tête au lieu d'une[27]). La bouche peut comporter des rangées de dents[26],[15],[30],[5],[32] ou un proboscis conique[33],[3]. Les yeux peuvent être représentés par une ou plusieurs[34] paires d'ocelles simples[3], comme on le voit chez Paucipodia[26], Luolishania[25], Miraluolishania (en)[34], Ovatiovermis[35], Onychodictyon[33], Hallucigenia[15] et peut-être aussi Aysheaia[33]. Chez les lobopodiens à branchies comme Kerygmachela, les yeux sont cependant des taches réflectrices relativement complexes[36],[37].
Tronc
[modifier | modifier le code]Le tronc est allongé et constitué de nombreux segments (métamères) porttant chacun une paire de lobopodes, mais la limite des segments n'a pas de signification externe comme chez les arthropodes. Ces segments peuvent porter d'autres structures comme des tubercules (par exemple chez Hadranax (en)[4] et Kerygmachela[11]), des sclérites épineuses ou en forme de plaques (par exemple chez les lobopodiens cuirassés[3]) ou des palettes natatoires (chez les lobopodiens à branchies, par exemple[11],[32]). La différentiation des appendices du tronc est rare, sauf chez les luolishanides et les hallucigenides, ou beaucoup de paires de lobopodes antérieurs sont nettement plus minces que celles de l'arrière[3],[35]. Le tronc peut se terminer par une paire de lobopodes (par exemple chez Aysheaia et Hallucigenia)[15] ou une extension ressemblant à une queue (par exemple chez Siberion et Jianshanopodia)[5],[38].
Structures internes
[modifier | modifier le code]
L'intestin des lobopodiens est souvent droit et indifférencié[39]. Certains spécimens ont l'intestion rempli de sédiments[26]. Cet intestin est constitué d'un tube central sur toute la longueur du tronc de l'animal[5], sans beaucoup varier en largeur — en tous cas pas systématiquement. Cependant dans certains groupes, spécifiquement les lobopodiens à branchies et les siberides, l'intestin est entouré par une série de paires de diverticules en forme de reins (des glandes digestives)[5],[30],[39]. Chez certains spécimens, une partie de l'intestin est parfois fossilisé en trois dimensions. Ceci ne peut pas provenir de la phosphatisation (en), habituellement responsable de ce phénomène[40] car le contenu en phosphate de l'intestin des lobopodiens est inférieur à 1% ; on y trouve du quartz et de la muscovite[26]. L'intestin de Paucipodia est plus large au milieu du tronc. Il est lâchement fixé dans la cavité corporelle et peut donc être flexible.

On ne sait pas grand-chose de l'anatomie nerveuse des lobopodiens en raison de la rareté et de l'ambiguïté des preuves fossiles. On a peut-être trouvé des traces de cerveau chez Megadictyon et de corde nerveuse ventrale (en) chez Paucipodia[30],[26]. La première existence avérée de structures nerveuses provient du lobopodien à branchies Kerygmachela dans une étude de 2018 — il présente un cerveau composé seulement d'un protocerebrum (en) (la partie la plus frontale du ganglion cérébral des panarthropodes) directement connecté aux nerfs des yeux et des appendices frontal, ce qui suggère l'origine protocérébrale de la tête des lobopodiens et des panarthropodes[36].
Chez certains ecdysozoaires actuels comme les priapulides et les onychophores, il existe une couche externes de muscles circulaires et une couche interne de muscles longitudinaux (les onychophores ont aussi une couche intermédiaire de muscles obliques entre les deux autres). Mais la musculature du lobopodien fossile Tritonychus (en) présente le schéma opposé : ce sont les muscles extérieurs qui sont longitudinaux et les muscles intérieurs sont circulaires[41].
Types de lobopodiens
[modifier | modifier le code]Les lobopodiens peuvent être classés en différents types selon leur morphologie externe — par exemple le taxon général vermiforme comme xenusides (en), les xenusides possédant des sclérites comme « lobopodiens cuirassés » et les taxons possédant à la fois des appendices frontaux robustes et des palettes latérales comme « lobopodiens à branchies ». Certains de ces types ont été définis à l'origine dans un sens taxonomiques (par exemple les xenusides comme la classe Xenusia), mais aucun d'eux n'est généralement accepté comme monophylétique par les études ultérieures[14],[16],[17].
Lobopodiens cuirassés
[modifier | modifier le code]

Les lobopodiens cuirassés sont les xenusides (en) porteurs de rangées de sclérites comme des épines ou des plaques le long de leur tronc (par exemple Hallucigenia, Microdictyon et Luolishania) ou sur leurs lobopodes (par exemple Diania). Par contraste, les lobopodiens sans sclérites sont parfois appelés « lobopodiens non-cuirassés »[27],[12]. Ces sclérites ont été interprétées comme ayant un rôle d'armure protectrice ou de points de fixation pour les muscles[42],[3]. Dans certains cas, seuls les sclérites désarticulées de l'animal ont été préservées au sein des petits fossiles coquilliers (SSF)[9]. Certaines études ont suggéré que les lobopodiens cuirassés étaient liés aux onychophores et pourraient même représenter un clade[42], mais leur position phylogénétique dans les études ultérieures est controversée (voir plus bas).
Lobopodiens à branchies
[modifier | modifier le code]
Les dinocaridides proches des lobopodiens (du fait de caractères communs comme les anneaux et les lobopodes) sont appelés « lobopodiens à branchies ». Ces formes présentent une paire de palettes sur chaque segment du tronc, mais pour le reste ne montrent aucun signe d'arthropodisation — par contraste avec les dinocaridides dérivés comme les Radiodonta (en) qui ont des appendices frontaux robustes et sclérotisés. Les lobopodiens à branchies comptent au moins deux genres : Pambdelurion et Kerygmachela[12]. Opabinia peut aussi être placé dans ce groupe au sens large[16],[43], bien que la présence de lobopodes dans ce genre ne soit pas définitivement prouvée[44]. Omnidens (en), un genre connu seulement par un appareil buccal proche de celui de Pambdelurion, pourrait aussi être un lobopodien à branchies[32]. Les palettes latérales pourraient avoir eu une double fonction d'appendices natatoires et de branchies[13] et sont peut-être homologues des palettes dorsales des radiodontes et des exopodes des arthropodes[11],[43]. La nature de lobopodiens de ces genres est encore contestée[45]. Ils sont cependant largement acceptés comme un groupe-racine des arthropodes basal des radiodontes[16],[14],[15],[17].
Siberion et les genres similaires
[modifier | modifier le code]
Siberion, Megadictyon et Jianshanopodia sont regroupés taxonomiquement par certaines études sous le nom de Siberiida, ou de siberidés[38]. Ce sont généralement de grands xenusides (longueur entre 7[38] et 22 centimètres[30]) au tronc large, aux lobopodes robustes sans trace de griffes et, de façon plus marquante, avec une paire de robustes appendices frontaux[16]. À l'exception de Siberion, ils ont aussi des glandes digestives comme celles des lobopodiens à branchies et des premiers euarthropodes[16],[39]. Leur anatomie représente des formes de transition entre les xénusides typiques et les lobopodiens à branchies[38], ce qui les place finalement dans la position la plus basale du groupe-racine des arthropodes[5],[30],[16],[17].
Paléoécologie
[modifier | modifier le code]
Les lobopodiens peuvent avoir occupé un grand nombre de niches écologiques. Bien que la plupart aient des appendices indifférenciés et un intestin droit, ce qui suggère un mode de vie de simples filtreurs de sédiments[3], les glandes digestives complexes et la grande taille des lobopodiens à branchies leur permettaient de consommer des aliments plus gros[3],[39] et leurs appendices frontaux robustes suggèrent même un mode de vie prédateur[5],[39]. D'un autre côté, les luolishaniides comme Luolishania et Ovatiovermis ont des lobopodes plumeux complexes qui formaient sans doute des « paniers » pour filtrer les particules en suspension[25],[35]. Les lobopodes terminés par des griffes courbes peuvent avoir donné à leurs possesseurs la capacité de grimper sur leur environnement[3].
On connait peu de choses de la physiologie des lobopodiens. Certains éléments suggèrent qu'ils muaient comme les autres ecdysozoaires, mais le contour et l'ornementation de leurs sclérites ne changeaient pas durant l'ontogenèse[10]. La structure semblable à des branchies des palettes des lobopodiens à branchies et les extensions ramifiées des lobopodes de Jianshanopodia peuvent avoir joué un rôle respiratoire (branchies)[13],[5]. Pambdelurion pourrait avoir contrôlé le mouvement de ses lobopodes d'une manière similaire à celle des onychophores[46].
Distribution
[modifier | modifier le code]Au Cambrien, les lobopodiens présentent un degré de biodiversité significatif. On connaît une espèce pour l'Ordovicien, une pour le Silurien[7],[47], et un peu plus au Carbonifère (Mazon Creek) — ce qui traduit la rareté des Lagerstätten exceptionnels dans les dépôts post-Cambrien.
Phylogénie
[modifier | modifier le code]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogenie neutralisée entre les lobopodiens et les autres taxons d'Ecdysozoa[14],[16],[15],[43],[17],[35],[18]. Les taxons de panarthropodes actuels sont en gras. Les relations entre leurs embranchements ne sont pas résolues. |




| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogénie d’après Pates et al., 2022 faisant des Lobopodiens un groupe paraphylétique basal d'Arthropodes. |
L'interprétation générale de la phylogénie des lobopodiens a changé radicalement au cours des décennies[3]. Cette réorganisation n'est pas seulement basée sur de nouvelles preuves fossiles, mais aussi sur de nouvelles informations embryologiques, neuroanatomiques et génomiques (expression génétique, phylogénomique) obtenues à partir de taxons de panarthropodes existants[3],[17],[48].
Les lobopodiens étaient traditionnellement considérés comme un groupe d'onychophores paléozoïques, sur la base de leur morphologie apparemment proche (par exemple leur cuticule annelée et leurs appendices lobopodes munis de griffes)[3],[49],[50],[51]. Cette interprétation a été remise en question après la découverte de lobopodiens avec des caractères d'arthropodes et de tardigrades[52], qui suggère que les similarités entre les lobopodiens et les onychophores sont des traits ancestraux des panarthropodes (plésiomorphies) plutôt que des traits exclusivement propres aux onychophores (synapomorphies)[18]. Le paléontologiste britannique Graham Budd considère par exemple les lobopodiens comme un niveau basal d'où seraient issus les embranchements des onychophores et des arthropodes, Aysheaia possédant un pan d'organisation comparable au plan d'organisation ancestral et des formes comme Kerygmachela et Pambdelurion représentant une transition conduisant à celui des arthropodes par l'intermédiaire des dinocaridides[42]. L'ornementation de surface d’Aysheaia, si elle est homologue des sclérites des paléoscolecides (en), peut représenter un lien qui le relie au groupe Cycloneuralia[42]. De nombreuses études ont suivi et développé cette idée, admettant généralement que les trois embranchements de panarthropodes ont des lobopodiens parmi leurs ancêtres[3],[14],[15],[17],[35],[18]. Les lobopodiens seraient donc paraphylétiques et comprendraient le dernier ancêtre commun des arthropodes, des onychophores et des tardigrades[3].
Comme groupe-racine des arthropodes
[modifier | modifier le code]Comparé aux autres groupes-racines des panarthropodes, la suggestion de membres lobopodiens du groupe-racine des arthropodes est relativement cohérente — des siberides comme Megadictyon et Jianshanopodia occupant la position la plus basale, puis les lobopodiens à branchies Pambdelurion et Kerygmachela et finalement un clade composé d’Opabinia, des radiodontes (en) et des euarthropodes (groupe-couronne des arthropodes)[14],[16],[15],[43],[17],[35],[18]. Leurs positions au sein du groupe-racine des arthropodes sont indiquées par de nombreux caractères de base communs et formes intermédiaires (par exemple des glandes digestives de type arthropode, des appendices frontaux de type radiodonte et des structures appendiculaires dorso-ventrales liées aux appendices biramés des arthropodes)[16],[17]. L'origine lobopodienne des arthropodes est aussi par des études génomiques des taxons actuels — l'expression génétique soutient l'homologie des appendices des arthropodes et des lobopodes des onychophores, ce qui suggère que les appendices segmentés modernes ont évolué à partir des lobopodes annelés, les appendices multi-segmentés des euarthropodes basaux éteints (par exemple les fuxianhuides (en)) constituant peut-être une forme intermédiaire[31]. D'autre part, les antennes primaires et les appendices frontaux des lobopodiens et des dinocaridides pourraient être homologues du complexe labrum/hypostome des euarthropodes, une idée renforcée par leur commune origine protocérébrale[16],[17],[36] et le mode de développement du labrum des arthropodes actuels[31],[17].
-
 Les radiodontes (en) sont un groupe-racine des arthropodes possédant des palettes corporelles, des appendices frontaux arthropodisés et des yeux composés sur pédoncule.
Les radiodontes (en) sont un groupe-racine des arthropodes possédant des palettes corporelles, des appendices frontaux arthropodisés et des yeux composés sur pédoncule. -
 Les membres multisegmentés des fuxianhuides (en) pourraient représenter une forme intermédiaire entre les lobopodes et les appendices des arthropodes modernes.
Les membres multisegmentés des fuxianhuides (en) pourraient représenter une forme intermédiaire entre les lobopodes et les appendices des arthropodes modernes.


Diania, un genre de lobopodiens cuirassés avec des pattes robustes et épineuses, était à l'origine associé au groupe-racine des arthropodes sur la base de ses appendices apparemment arthropodisés[29]. Cette interprétation est cependant questionnable, dans la mesure où les données de la description originale ne sont pas cohérentes avec les relations phylogénétiques supposées[53],[54]. Un réexamen plus tardif a même révélé que l'arthropodisation supposée des pattes de Diania était une mauvaise interprétation — bien que leurs épines puissent avoir été dures, le reste de la cuticule des pattes de Diania était mou (ni durci, ni sclérotisé), sans aucune trace de pivot ni de membrane arthrodiale, ce qui suggère que ces pattes sont des lobopodes avec seulement des anneaux largement espacés[28],[12]. Le réexamen rejette donc finalement les preuves d'arthropodisation des appendices (sclérotisation, segmentation et articulation), ainsi que la relation fondamentale entre Diania et les arthropodes[28],[12].
Comme groupe-racine des onychophores
[modifier | modifier le code]
Si Antennacanthopodia est largement accepté comme membre du groupe-racine des onychophores[14],[15],[17],[35],[18] la position d'autres genres de xenusides auparavant considérés comme liés aux onychophores est controversée — des études plus récentes suggèrent qu'ils appartiennent, soit au groupe-racine des onychophores[3],[14],[17], soit aux panarthropodes basaux[35],[18], avec quelques espèces occasionnellement proposées comme appartenant au groupe-racine des tardigrades et/ou à celui des panarthropodes[35],[18]. Une étude de 2014 suggère qu’Hallucigenia fait partie du groupe-racine des onychophores, sur la base de ses griffes, dont les structures internes superposées ressemblent à celles d'un onychophore actuel[14]. Cette interprétation a été remise en cause par des études plus récentes, ces structures pouvant représenter un trait ancestral des panarthropodes[18].
Comme groupe-racine des tardigrades
[modifier | modifier le code]Les taxons lobobodiens du groupe-racine des tardigrades ne sont pas clairs[3]. Aysheaia[35],[18] ou Onychodictyon ferox[14],[15] ont été suggérés comme membres possibles. Il y a même eu des suggestions (pas généralement acceptées) que Tardigrada lui-même représente la branche la plus basale des panarthropodes, ou se rattache au groupe-racine des arthropodes[52].
Comme groupe-racine des panarthropodes
[modifier | modifier le code]On ne sait pas bien quels lobopodiens représentent des membres du groupe-racine des panarthropodes, ni lesquels se branchent juste avant le dernier ancêtre commun des embranchements de panarthropodes actuels. Aysheaia pourrait avoir occupé cette position, si l'on en juge par sa morphologie apparemment basale[42],[14],[15], mais d'autres études suggèrent plutôt les luolishanides et les hallucigenides[35],[18], deux taxons de lobopodiens qui ont été aussi classés comme membres du groupe-racine des onychophores[3],[14],[17].
Genres décrits
[modifier | modifier le code]-
 Fossile d’Aysheaia pedunculata.
Fossile d’Aysheaia pedunculata. -
 Fossile de Microdictyon sp.
Fossile de Microdictyon sp. -
 Fossile de Mureropodia apae, qui est peut-être en fait l'appendice frontal de Caryosyntrips (en) camurus.
Fossile de Mureropodia apae, qui est peut-être en fait l'appendice frontal de Caryosyntrips (en) camurus. -
 Reconstitution de Facivermis, un lobopodien inhabituel sans lobopodes postérieurs.
Reconstitution de Facivermis, un lobopodien inhabituel sans lobopodes postérieurs.



En 2018, plus de 20 genres de lobopodiens avaient été décrits[12]. Les fossiles décrits sous les noms de Mureropodia (en) apae et Aysheaia prolata sont respectivement considérés comme des appendices frontaux désarticulés des radiodontes (en) Caryosyntrips (en) et Stanleycaris[55],[56],[57]. Certains auteurs ont suggéré que Miraluolishania (en) est synonyme de Luolishania[58],[59]. L'énigmatique Facivermis s'est révélé un genre de lobopodien luolishanide hautement spécialisé[38],[35],[60].
- Antennacanthopodia
- Aysheaia
- Carbotubulus
- Cardiodictyon (en)
- Collinsium (en)
- Collinsovermis[61]
- Diania
- Facivermis
- Hadranax (en)
- Hallucigenia
- Jianshanopodia
- Kerygmachela
- Lenisambulatrix
- Luolishania
- Megadictyon
- Microdictyon
- Miraluolishania (en) (= Luolishania ?)
- Onychodictyon
- Orstenotubulus (en)
- Ovatiovermis
- Pambdelurion
- Paucipodia
- Siberion
- Thanahita
- Tritonychus (en)
- Xenusion (en)
Liens externes
[modifier | modifier le code]- Ressource relative au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :
Notes et références
[modifier | modifier le code]Notes
[modifier | modifier le code]Références
[modifier | modifier le code]- (en) Snodgrass, R.E., « Evolution of the Annelida, Onychophora, and Arthropoda », Smithsonian Miscellaneous Collections, vol. 97, no 6, , p. 1–159 (lire en ligne)
- (en) T. Cavalier-Smith, « A revised six-kingdom system of life », Biological Reviews, vol. 73, no 3, , p. 203–266 (PMID 9809012, DOI 10.1017/S0006323198005167)
- (en) Javier Ortega-Hernández, « Lobopodians », Current Biology, vol. 25, no 19, , R873–R875 (ISSN 0960-9822, PMID 26439350, DOI 10.1016/j.cub.2015.07.028, lire en ligne
 )
)
- (en) Graham Budd et John Peel, « A new Xenusiid lobopod from the early Cambrian Sirius Passet fauna of North Greenland », Palaeontology, vol. 41, , p. 1201–1213 (lire en ligne)
- (en) Jianni Liu, Degan Shu, Jian Han, Zhifei Zhang et Xingliang Zhang, « A large xenusiid lobopod with complex appendages from the Lower Cambrian Chengjiang Lagerstätte », Acta Palaeontol. Pol., vol. 51, no 2, , p. 215–222 (lire en ligne, consulté le )
- (en) P. van Roy, P.J. Orr, J.P. Botting, L.A. Muir, J. Vinther, B. Lefebvre, K.E. Hariri et D.E.G. Briggs, « Ordovician faunas of Burgess Shale type », Nature, vol. 465, no 7295, , p. 215–218 (PMID 20463737, DOI 10.1038/nature09038, Bibcode 2010Natur.465..215V, S2CID 4313285)
- (en) P.H. von Bitter, M.A. Purnell, D.K. Tetreault et C.A. Stott, « Eramosa Lagerstätte—Exceptionally preserved soft-bodied biotas with shallow-marine shelly and bioturbating organisms (Silurian, Ontario, Canada) », Geology, vol. 35, no 10, , p. 879 (DOI 10.1130/G23894A.1, Bibcode 2007Geo....35..879V, S2CID 11561169, lire en ligne)
- (en) J.T. Haug, G. Mayer, C. Haug et D.E.G. Briggs, « A Carboniferous non-Onychophoran Lobopodian reveals long-term survival of a Cambrian morphotype », Current Biology, vol. 22, no 18, , p. 1673–1675 (PMID 22885062, DOI 10.1016/j.cub.2012.06.066)
- (en) J.-B. Caron, M.R. Smith et T.H.P. Harvey, « Beyond the Burgess Shale: Cambrian microfossils track the rise and fall of hallucigeniid lobopodians. », Proceedings of the Royal Society B: Biological Sciences, vol. 280, no 1767, , p. 20131613 (PMID 23902914, PMCID 3735267, DOI 10.1098/rspb.2013.1613)
- (en) T.P. Topper, C.B. Skovsted, J.S. Peel et D.A.T. Harper, « Moulting in the lobopodian Onychodictyonfrom the lower Cambrian of Greenland », Lethaia, , n/a (DOI 10.1111/let.12026, lire en ligne)
- (en) Graham E. Budd, « The morphology and phylogenetic significance of Kerygmachela kierkegaardi Budd (Buen Formation, Lower Cambrian, N Greenland) », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 89, no 4, , p. 249–290 (ISSN 1473-7116, DOI 10.1017/S0263593300002418)
- (en) Qiang Ou et Georg Mayer, « A Cambrian unarmoured lobopodian, †Lenisambulatrix humboldti gen. et sp. nov., compared with new material of †Diania cactiformis », Scientific Reports, vol. 8, no 1, , p. 13667 (PMID 30237414, PMCID 6147921, DOI 10.1038/s41598-018-31499-y, Bibcode 2018NatSR...813667O, lire en ligne )
- (en) Graham Budd, « A Cambrian gilled lobopod from Greenland », Nature, vol. 364, no 6439, , p. 709–711 (ISSN 1476-4687, DOI 10.1038/364709a0, Bibcode 1993Natur.364..709B, S2CID 4341971)
- (en) Martin R. Smith et Javier Ortega-Hernández, « Hallucigenia's onychophoran-like claws and the case for Tactopoda », Nature, vol. 514, no 7522, , p. 363–366 (PMID 25132546, DOI 10.1038/nature13576, Bibcode 2014Natur.514..363S, S2CID 205239797, lire en ligne)
- (en) Martin R. Smith et Jean-Bernard Caron, « Hallucigenia's head and the pharyngeal armature of early ecdysozoans », Nature, vol. 523, no 7558, , p. 75–78 (ISSN 0028-0836, PMID 26106857, DOI 10.1038/nature14573, Bibcode 2015Natur.523...75S, S2CID 205244325, lire en ligne)
- (en) Javier Ortega-Hernández, « Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848 », Biological Reviews of the Cambridge Philosophical Society, vol. 91, no 1, , p. 255–273 (ISSN 1469-185X, PMID 25528950, DOI 10.1111/brv.12168, S2CID 7751936)
- (en) Javier Ortega-Hernández, Ralf Janssen et Graham E. Budd, « Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective », Arthropod Structure & Development, vol. 46, no 3, , p. 354–379 (ISSN 1467-8039, PMID 27989966, DOI 10.1016/j.asd.2016.10.011)
- (en) Derek J. Siveter, Derek E. G. Briggs, David J. Siveter, Mark D. Sutton et David Legg, « A three-dimensionally preserved lobopodian from the Herefordshire (Silurian) Lagerstätte, UK », Royal Society Open Science, vol. 5, no 8, , p. 172101 (PMID 30224988, PMCID 6124121, DOI 10.1098/rsos.172101)
- (en) Jerzy Dzik et Günter Krumbiegel, « The oldest 'onychophoran' Xenusion: a link connecting phyla? », Lethaia, vol. 22, no 2, , p. 169–181 (ISSN 1502-3931, DOI 10.1111/j.1502-3931.1989.tb01679.x)
- (en) Frank W. Smith et Bob Goldstein, « Segmentation in Tardigrada and diversification of segmental patterns in Panarthropoda », Arthropod Structure & Development, vol. 46, no 3, , p. 328–340 (ISSN 1467-8039, PMID 27725256, DOI 10.1016/j.asd.2016.10.005)
- (en) Pentastomida - Sociedad Entomológica Aragonesa
- (en) Treatise on Zoology - Anatomy, Taxonomy, Biology. The Crustacea, Volum 5
- (en) Richard A. Fortey et Richard H. Thomas, Arthropod Relationships, Springer Science & Business Media, (ISBN 978-0-412-75420-3, lire en ligne)
- (en) Gould, S.J., Wonderful Life: The Burgess Shale and the Nature of History, W.W. Norton & Company, (Bibcode 1989wlbs.book.....G)[page à préciser]
- (en) Xiaoya Ma, Xianguang Hou et Jan Bergström, « Morphology of Luolishania longicruris (Lower Cambrian, Chengjiang Lagerstätte, SW China) and the phylogenetic relationships within lobopodians », Arthropod Structure & Development, vol. 38, no 4, , p. 271–291 (ISSN 1467-8039, PMID 19293001, DOI 10.1016/j.asd.2009.03.001, lire en ligne)
- (en) Hou, Xian-Guang, Xiao-Ya Ma, Jie Zhao et Jan Bergström, « The lobopodian Paucipodia inermis from the Lower Cambrian Chengjiang fauna, Yunnan, China », Lethaia, vol. 37, no 3, , p. 235–244 (DOI 10.1080/00241160410006555)
- (en) Qiang Ou, Jianni Liu, DEGAN SHU, Jian Han, Zhifei Zhang, Xiaoqiao Wan et Qianping Lei, « A Rare Onychophoran-Like Lobopodian from the Lower Cambrian Chengjiang Lagerstätte, Southwestern China, and its Phylogenetic Implications », Journal of Paleontology, vol. 85, no 3, , p. 587–594 (DOI 10.1666/09-147R2.1, JSTOR 23020193, S2CID 53056128, lire en ligne)
- (en) Xiaoya Ma, Gregory Edgecombe, David Legg et Xianguang Hou, « The morphology and phylogenetic position of the Cambrian lobopodian Diania cactiformis », Journal of Systematic Palaeontology, vol. 12, no 4, , p. 445–457 (DOI 10.1080/14772019.2013.770418, S2CID 220463025, lire en ligne)
- (en) Jianni Liu, Michael Steiner, Jason A. Dunlop, Helmut Keupp, Degan Shu, Qiang Ou, Jian Han, Zhifei Zhang et Xingliang Zhang, « An armoured Cambrian lobopodian from China with arthropod-like appendages », Nature, vol. 470, no 7335, , p. 526–530 (ISSN 1476-4687, PMID 21350485, DOI 10.1038/nature09704, Bibcode 2011Natur.470..526L, S2CID 4324509)
- (en) J. Liu, D. Shu, J. Han, Z. Zhang et X. Zhang, « Morpho-anatomy of the lobopod Magadictyon cf. Haikouensis from the Early Cambrian Chengjiang Lagerstätte, South China », Acta Zoologica, vol. 88, no 4, , p. 279–288 (DOI 10.1111/j.1463-6395.2007.00281.x)
- (en) Elizabeth L. Jockusch, « Developmental and Evolutionary Perspectives on the Origin and Diversification of Arthropod Appendages », Integrative and Comparative Biology, vol. 57, no 3, , p. 533–545 (ISSN 1540-7063, PMID 28957524, DOI 10.1093/icb/icx063)
- (en) Jakob Vinther, Luis Porras, Fletcher Young, Graham Budd et Gregory Edgecombe, « The mouth apparatus of the Cambrian gilled lobopodian Pambdelurion whittingtoni », Palaeontology, vol. 59, no 6, , p. 841–849 (DOI 10.1111/pala.12256, lire en ligne)
- (en) Qiang Ou, Degan Shu et Georg Mayer, « Cambrian lobopodians and extant onychophorans provide new insights into early cephalization in Panarthropoda », Nature Communications, vol. 3, no 1, , p. 1261 (ISSN 2041-1723, PMID 23232391, PMCID 3535342, DOI 10.1038/ncomms2272, Bibcode 2012NatCo...3.1261O)
- (en) Brigitte Schoenemann, Jian-Ni Liu, De-Gan Shu, Jian Han et Zhi-Fei Zhang, « A miniscule optimized visual system in the Lower Cambrian », Lethaia, vol. 42, no 3, , p. 265–273 (ISSN 1502-3931, DOI 10.1111/j.1502-3931.2008.00138.x)
- (en) Jean-Bernard Caron et Cédric Aria, « Cambrian suspension-feeding lobopodians and the early radiation of panarthropods », BMC Evolutionary Biology, vol. 17, no 1, , p. 29 (ISSN 1471-2148, PMID 28137244, PMCID 5282736, DOI 10.1186/s12862-016-0858-y)
- (en) Tae-Yoon S. Park, Ji-Hoon Kihm, Jusun Woo, Changkun Park, Won Young Lee, M. Paul Smith, David A. T. Harper, Fletcher Young et Arne T. Nielsen, « Brain and eyes of Kerygmachela reveal protocerebral ancestry of the panarthropod head », Nature Communications, vol. 9, no 1, , p. 1019 (ISSN 2041-1723, PMID 29523785, PMCID 5844904, DOI 10.1038/s41467-018-03464-w, Bibcode 2018NatCo...9.1019P)
- (en) James F. Fleming, Reinhardt Møbjerg Kristensen, Martin Vinther Sørensen, Tae-Yoon S. Park, Kazuharu Arakawa, Mark Blaxter, Lorena Rebecchi, Roberto Guidetti, Tom A. Williams, Nicholas W. Roberts et Jakob Vinther, « Molecular palaeontology illuminates the evolution of ecdysozoan vision », Proceedings of the Royal Society B: Biological Sciences, vol. 285, no 1892, , p. 20182180 (ISSN 0962-8452, PMID 30518575, PMCID 6283943, DOI 10.1098/rspb.2018.2180)
- (en) Jerzy Dzik, « The xenusian-to-anomalocaridid transition within the lobopodians », Bollettino della Societa Paleontologica Italiana, vol. 50, , p. 65–74 (lire en ligne)
- (en) Jean Vannier, Jianni Liu, Rudy Lerosey-Aubril, Jakob Vinther et Allison C. Daley, « Sophisticated digestive systems in early arthropods », Nature Communications, vol. 5, no 1, , p. 3641 (ISSN 2041-1723, PMID 24785191, DOI 10.1038/ncomms4641, Bibcode 2014NatCo...5.3641V)
- (en) N. J. Butterfield, « Leanchoilia guts and the interpretation of three-dimensional structures in Burgess Shale-type fossils », Paleobiology, vol. 28, , p. 155–171 (ISSN 0094-8373, DOI 10.1666/0094-8373(2002)028<0155:LGATIO>2.0.CO;2)
- (en) Zhang X-G, Smith MR, YangJ, Hou J-B. 2016 Onychophoran-like musculature in a phosphatized Cambrian lobopodian Biol. Lett. 12: 20160492. https://dx.doi.org/10.1098/rsbl.2016.0492 (consulté le 25 janvier 2021)
- (en) G. E. Budd, « Why are arthropods segmented? », Evolution and Development, vol. 3, no 5, , p. 332–42 (PMID 11710765, DOI 10.1046/j.1525-142X.2001.01041.x, S2CID 37935884)
- (en) Peter Van Roy, Allison C. Daley et Derek E. G. Briggs, « Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps », Nature, vol. 522, no 7554, , p. 77–80 (ISSN 1476-4687, PMID 25762145, DOI 10.1038/nature14256, Bibcode 2015Natur.522...77V, S2CID 205242881)
- (en) Graham E. Budd et Allison C. Daley, « The lobes and lobopods of Opabinia regalis from the middle Cambrian Burgess Shale: The lobes of Opabinia », Lethaia, vol. 45, no 1, , p. 83–95 (DOI 10.1111/j.1502-3931.2011.00264.x, lire en ligne)
- (en) J.-y Chen, L. Ramskold et G.-q Zhou, « Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators », Science, vol. 264, no 5163, , p. 1304–1308 (ISSN 0036-8075, PMID 17780848, DOI 10.1126/science.264.5163.1304, Bibcode 1994Sci...264.1304C, S2CID 1913482, lire en ligne)
- (en) Fletcher J. Young et Jakob Vinther, « Onychophoran-like myoanatomy of the Cambrian gilled lobopodian Pambdelurion whittingtoni », Palaeontology, vol. 60, no 1, , p. 27–54 (ISSN 1475-4983, DOI 10.1111/pala.12269, lire en ligne)
- (en) R. J. Whittle, S. E. Gabbott, R. J. Aldridge et J. Theron, « An Ordovician Lobopodian from the Soom Shale Lagerstätte, South Africa », Palaeontology, vol. 52, no 3, , p. 561–567 (DOI 10.1111/j.1475-4983.2009.00860.x)
- (en) Gregory D. Edgecombe, « Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa », Evolution: Education and Outreach, vol. 2, no 2, , p. 178–190 (ISSN 1936-6434, DOI 10.1007/s12052-009-0118-3)
- (en) L. Ramsköld et Hou Xianguang, « New early Cambrian animal and onychophoran affinities of enigmatic metazoans », Nature, vol. 351, no 6323, , p. 225–228 (ISSN 1476-4687, DOI 10.1038/351225a0, Bibcode 1991Natur.351..225R, S2CID 4309565)
- (en) J. Bergström et Xian-Guang Hou, « Cambrian Onychophora or Xenusians », Zoologischer Anzeiger - ZOOL ANZ, vol. 240, nos 3–4, , p. 237–245 (DOI 10.1078/0044-5231-00031, lire en ligne)
- (en) R. A. Robison, « Affinities of Aysheaia (Onychophora), with Description of a New Cambrian Species », Journal of Paleontology, vol. 59, no 1, , p. 226–235 (ISSN 0022-3360, JSTOR 1304837)
- (en) Graham E. Budd, « Tardigrades as 'Stem-Group Arthropods': The Evidence from the Cambrian Fauna », Zoologischer Anzeiger - A Journal of Comparative Zoology, vol. 240, no 3, , p. 265–279 (ISSN 0044-5231, DOI 10.1078/0044-5231-00034)
- (en) Ross C. P. Mounce et Matthew A. Wills, « Phylogenetic position of Diania challenged », Nature, vol. 476, no 7359, , E1; discussion E3–4 (ISSN 1476-4687, PMID 21833044, DOI 10.1038/nature10266, Bibcode 2011Natur.476E...1M, S2CID 4417903, lire en ligne)
- (en) David Legg, Xiaoya Ma, Joanna Wolfe, Javier Ortega-Hernández, Gregory Edgecombe et Mark Sutton, « Lobopodian phylogeny reanalysed », Nature, vol. 476, no 7359, , E2–3; discussion E3 (PMID 21833046, DOI 10.1038/nature10267, Bibcode 2011Natur.476Q...1L, S2CID 4310063, lire en ligne)
- (en) Stephen Pates et Allison C. Daley, « Caryosyntrips: a radiodontan from the Cambrian of Spain, USA and Canada », Papers in Palaeontology, vol. 3, no 3, , p. 461–470 (ISSN 2056-2802, DOI 10.1002/spp2.1084, lire en ligne)
- (en) « Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris - Acta Palaeontologica Polonica », sur www.app.pan.pl (consulté le )
- (en) « Reply to Comment on "Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris" with the formal description of Stanleycaris - Acta Palaeontologica Polonica », sur www.app.pan.pl (consulté le )
- (en) Xiaoya Ma, Xianguang Hou et Jan Bergström, « Morphology of Luolishania longicruris (Lower Cambrian, Chengjiang Lagerstätte, SW China) and the phylogenetic relationships within lobopodians », Arthropod Structure & Development, vol. 38, no 4, , p. 271–291 (ISSN 1467-8039, PMID 19293001, DOI 10.1016/j.asd.2009.03.001)
- (en) Jianni Liu, DeGan Shu, Jian Han et Zhifei Zhang, « Comparative study of Cambrian lobopods Miraluolishania and Luolishania », Chinese Science Bulletin - CHIN SCI BULL, vol. 53, no 1, , p. 87–93 (DOI 10.1007/s11434-007-0428-1, Bibcode 2008ChSBu..53...87L, S2CID 128619311, lire en ligne)
- (en) Richard J. Howard, Xianguang Hou, Gregory D. Edgecombe, Tobias Salge, Xiaomei Shi et Xiaoya Ma, « A Tube-Dwelling Early Cambrian Lobopodian », Current Biology, vol. 0, no 8, , p. 1529–1536.e2 (ISSN 0960-9822, PMID 32109391, DOI 10.1016/j.cub.2020.01.075, lire en ligne )
- (en) Jean-Bernard Caron et Cédric Aria, « The Collins' monster, a spinous suspension-feeding lobopodian from the Cambrian Burgess Shale of British Columbia », Palaeontology, vol. n/a, no n/a, (ISSN 1475-4983, DOI 10.1111/pala.12499, lire en ligne)